| Citation: | M Ben Fadhel, T Zouaghi, A Amri, M Ben Youssef. Radiolarian and Planktic Foraminifera Biostratigraphy of the Early Albian Organic Rich Beds of Fahdene Formation, Northern Tunisia. Journal of Earth Science, 2014, 25(1): 45-63. doi: 10.1007/s12583-014-0399-5

|

| Amédro, F., 2008. Support for a Vraconnian Stage between the Albian Sensu Stricto and the Cenomanian (Cretaceous System). Carnet de Géologie, Memoire 2008/02(CG2008_M02) |
| Arthur, M. A., Sageman, B. B., 1994. Marine Black Shales: Depositional Mechanisms and Environments of Ancient Deposits. Annu. Rev. Earth Planet. Sci. , 22: 499–551 doi: 10.1146/annurev.ea.22.050194.002435 |
| Babazadeh, S. A., De Wever, P., 2004. Radiolarian Cretaceous Age of Soulabest Radiolarites in Ophiolite Suite of Eastern Iran. Bull. Soc. Géol. de France, 175(2): 121–129 doi: 10.2113/175.2.121 |
| Bak, M., 1995. Mid Cretaceous Radiolarians from the Pieniny Klippen Belt, Carpathians. Poland. Cretaceous Research, 16(1): 1–23 doi: 10.1006/cres.1995.1001 |
| Bak, M., 1999. Cretaceous Radiolarian Zonation in the Polish Part of the Pienny Klippen Belt (Western Carpathians). Geologica Carpathica, 50(1): 21–31 http://www.researchgate.net/profile/Marta_Bk/publication/271327667_Integrated_microbiostratigraphy_in_the_Maastrichtian_to_Paleocene_distal-flysch_sediments_of_the_Uzgrun_section_(Raa_unit_Carpathians_flysch_Czech_Republic)/links/55916cdc08ae15962d8e1f4b.pdf |
| Bak, M., 2004. Radiolarian Biostratigraphy of the Upper Cenomanian-Lower Turonian Deposits in the Subsilesian Nappe (Outer Western Carpathians). Geologica Carpathica, 55(3): 239–250 http://www.researchgate.net/profile/Marta_Bk/publication/271384553_Radiolarian_biostratigraphy_of_the_Upper_Cenomanian-Lower_Turonian_deposits_in_the_Subsilesian_Nappe_(Outer_Western_Carpathians)/links/54c67b380cf219bbe4f863b5.pdf |
| Bellier, J. P., Moullade, M., 2002. Lower Cretaceous Planktonic Foraminiferal Biostratigraphy of the Western North Atlantic (ODP LEG 171B), and Taxonomic Clarification of Key Index Species. Rev. de Micropal. , 45(1): 9–26 doi: 10.1016/S0035-1598(02)80003-4 |
| Bellier, J. P., Moullade, M., Huber, B. T., 2000. Mid-Cretaceous Planktic Foraminifers from Blake Nose: Revised Biostratigraphic Framework. In: Norris, R. D., Kroon, D., Klaus, A., eds., Proceedings of the Ocean Drilling Program, Scientific Results, 171B. Ocean Drilling Program, College Station, Texas |
| Ben Chelbi, M., Melki, F., Zargouni, F., 2006. Mode of Salt Bodies Emplacement in Septentrional Atlas of Tunisia: Example of a Bir Afou Salt Body. Comptes Rendus Géosciences, 338(5): 349–358, doi: 10.1016/j.crte.2006.02.009 |
| Ben Fadhel, M., Layeb, M., Ben Youssef, M., 2010. Upper Albian Planktic Foraminifera and Radiolarian Biostratigraphy (Nebeur-Northern Tunisia). Comptes Rendus Palevol, 9(3): 73–81 doi: 10.1016/j.crpv.2010.01.002 |
| Ben Fadhel, M., Layeb, M., Hedfi, A., et al., 2011. Albian Oceanic Anoxic Events in Northern Tunisia: Biostratigraphic and Geochemical Insights. Cretaceous Research, 32(6): 685–699 http://www.onacademic.com/detail/journal_1000035371792310_e14a.html |
| Ben Haj Ali, N., 2005. Les Foraminifères Planctoniques du Crétacé (Hauterivien à Turonien Inférieur) du Tunisie: Systématiques, Biozonations et Précisions Biostratigraphique: [Dissertation]. Université Tunis el Manar, Faculté des Sciences de Tunis, Tunis (in French) |
| Beurrier, M., Bourdillon-De-Grissac, C., De Wever, P., et al., 1987. Biostratigraphie des Radiolarites Associées Aux Volcanites Ophiolitiques de la Nappe de Samail (Sultanat d'Oman): Conséquences Tectogénétiques. Comp. Ren. Acad. Sci. , 304: 907–910 |
| Biely, A., Memmi, L., Salaj, J., 1973. Le Crétacé Inferieur de la Région d'Enfidaville. Decouverte d'Aptien Condense. Livr. Jub. M. Solignac, Ann. Min. Geol. , 26: 169–l78 |
| Bismuth, H., 1973. Réflexions Stratigraphiques Sur l'Albo-Aptien Dans la Région des Djebels Douleb et Semmama et Son Environnement (Tunisie du Centre-Nord). Ann. Min. Géol. , 26: 179–212 |
| Bismuth, H., Boltenhagen, C., Donze, P., et al., 1981. Le Crétacé Moyen et Supérieur du Djebel Semmama (Tunisie du Centre Nord), Microstratigraphie et évolution Sédimentologique. Bull. Cent. Rech. Expl. Prod. Elf-Aquit. , 5(11): 193–267 http://www.researchgate.net/publication/279617660_Middle_and_Upper_Cretaceous_in_the_Djebel_Semmama_northern_central_Tunisia_microstratigraphy_and_sedimentological_evolution_Le_Cretace_moyen_et_superieur_du_Djebel_Semmama_Tunisie_du_Centre-Nord_micro |
| Boltenhagen, C., 1985. Paléogéographie du Crétacé Moyen de la Tunisie Centrale. Proceedings of 1er Congrès Nat. Sc. Terre, Tunis. 97–114 |
| Boughdiri, M., Cordey, F., Sallouhi, H., et al., 2007. Jurassic Radiolarian-Bearing Series of Tunisia: Biostratigraphy and Significance to Western Tethys Correlations. Swiss Journal of Geosciences, 100(3): 431–441 doi: 10.1007/s00015-007-1237-x |
| Bralower, T. J., Sliter, W. V., Arthur, M. A., et al., 1993. Dysoxic/Anoxic Episodes in the Aptian-Albian (Early Cretaceous). In: Pringle, M. S., Sager, W. W., Sliter, W. V., et al., eds., The Mesozoic Pacific: Geology, Tectonics and Volcanism. Geo physical Monograph, 77: 5–37 |
| Bréhéret, J. G., 1988. Épisodes de Sédimentation Riche en Matière Organique Dans les Marnes Bleues d'âge Aptien et Albien de La Partie Pélagique du Bassin Vocontien. Bull. Soc. Géol. France, IV(2): 349–356 |
| Bréhéret, J. G., 1997. L'Aptien et l'Albien de la Fosse Vocontienne. Évolution de la Sédimentation et Enseignements sur les évènements Anoxiques. Soc. Géol. Nord, Publication, Villeneuve d'Ascq, 25: 614 |
| Bréhéret, J. G., Caron, M., Delamette, M., 1986. Niveaux Riches en Matière Organique Dans l'Albien Vocontien, Quelques Caractères du Paléoenviron-Nement, Essai d'interprétation Génétique. In: Bréhéret, J. G., ed., Les CouchesRiches en Matière Organique et Leurs Conditions de Dépôt. Documents du Bureau de Recherches Géologiques et Minières, Orléans, 110: 41–191 |
| Burollet, P. F., 1956. Contribution à l'étude Stratigraphique de la Tunisie Centrale. Annales des Mines et de Géologie, 18(1): 350 http://www.researchgate.net/publication/303602546_Contribution_a_l'etude_stratigraphique_de_la_Tunisie_centrale |
| Burollet, P. F., 1988. Cretaceous Reservoirs in Tunisia. AAPG Bull. , 72: 8 |
| Burollet, P. F., Memmi, L., M'Rabet, A., 1983. Le Crétacé Inférieur de Tunisie. Aperçu Stratigraphique et Sédimentologique. Zitteliana, 10: 255–264 |
| Chihaoui, A., 2009. La Transgression Albienne Dans la Région de Tajerouine en Tunisie Centrale: Stratigraphie, Sédimentologie et Tectonique Synsédimentaire: [Dissertation]. l'Université Joseph Fourier-Grenoble I, France |
| Chihaoui, A., Jaillard, E., Latil, J. L., et al., 2010. Stratigraphy of the Hameima and Lower Fahdene Formations in the Tadjerouine Area (Northern Tunisia). J. Afri. Earth Sciences, 58(2): 387–399 doi: 10.1016/j.jafrearsci.2010.02.008 |
| Cordey, F., Boughdiri, M., Sallouhi, H., 2005. First Direct Age Determination from the Jurassic Radiolarian-Bearing Siliceous Series (Jedidi Formation) of North-Western Tunisia. Comp. Rend. Geosc. , 337: 777–785 doi: 10.1016/j.crte.2005.03.013 |
| Dall'Agnolo, S., 2000. Le Crétacé de la Nappe de la Brèche (Préalpes Franco-Suisses). Données Nouvelles et Essai de Synthèse Stratigraphique et Paléogéographique. Eclog. Geol. Helve. , 93: 157–174 http://www.researchgate.net/publication/288394120_Le_cretace_de_la_nappe_de_la_breche_Prealpes_franco-suisses_Donnees_nouvelles_et_essai_de_synthese_stratigraphique_et_paleogeographique |
| Danelian, T., 2008. Diversity and Biotic Changes of Archaeodictyomitrid Radiolaria from the Aptian/Albian Transition (OAE1b) of Southern Albania. Micropaleontology, 54(1): 3–12 http://search.ebscohost.com/login.aspx?direct=true&db=aph&AN=32922077&site=ehost-live |
| Danelian, T., Baudin, F., Gardin, S., et al., 2007. The Record of Mid Cretaceous Oceanic Anoxic Events from the Ionian Zone of Southern Albania. Rev. Micropal. , 50(3): 225–237 doi: 10.1016/j.revmic.2007.06.004 |
| Danelian, T., Tsikos, H., Gardin, S., et al., 2004. Global and Regional Palaeoceanographic Changes as Recorded in the Mid-Cretaceous (Aptian-Albian) Sequence of the Ionian Zone (Northwestern Greece). J. Geol. Soc. , 161(6): 703–709 http://search.ebscohost.com/login.aspx?direct=true&db=aph&AN=13875258&site=ehost-live |
| de Wever, P., Baudin, F., 1996. Palaeogeography of Radiolarite and Organic-Rich Deposits in Mesozoic Tethys. Geologische Rundschau, 85(2): 310–326 doi: 10.1007/BF02422237 |
| Erba, E., Channell, J. E. T., Claps, M., et al., 1999. Integrated Stratigraphy of the Cismon Apticore (Southern Alps, Italy): A "Reference Section" for the Barremian-Aptian Interval at Low Latitudes. J. Foram. Res. , 29(4): 371–391 http://www.researchgate.net/publication/285020357_Integrated_stratigraphy_of_the_Cismon_APTICORE_Southern_Alps_Italy_A_Reference_section_for_the_Barremian-Aptian_interval_at_low_latitudes |
| Erbacher, J., 1998. Mid-Cretaceous Radiolarians from the Eastern Equatorial Atlantic and Their Paleoceanography. In: Mascle, J., Lohmann, G. P., Moullade, M., eds., Proceedings of the Ocean Program, Scientific Results, 159: 363–373 |
| Erbacher, J., Gerth, W., Schmiedl, G., et al., 1998. Benthic Foraminiferal Assemblages of Late Aptian-Early Albian Black Shale Intervals in the Vocontian Basin, SE France. Cretaceous Research, 19(6): 805–826 doi: 10.1006/cres.1998.0134 |
| Erbacher, J., Thurow, J., 1997. Influence of Oceanic Anoxic Events on the Evolution of Mid-Cretaceous Radiolaria in the North Atlantic and Western Tethys. Mar. Micropal. , 30(1–3): 139–158 http://eurekamag.com/pdf.php?pdf=008868610 |
| Erbacher, J., Thurow, J., 1998. Mid-Cretaceous Radiolarian Zonation for the North Atlantic: An Example of Oceanographically Controlled Evolutionary Processes in the Marine Biosphere? In: Cramp, A., Macleod, C. J., Lee, S. V., et al., eds., Geological Evolution of Ocean Basins: Results from Ocean Drilling Program. Geolo. Soc., London, Special Publications, 131(384): 71–82 |
| Fiet, N., Beaudoin, B., Parize, O., 2001. Lithostratigraphic Analysis of Milankovitch Cyclicity in Pelagic Albian Deposits of Central Italy: Implications for the Duration of the Stage and Substages. Cretaceous Research, 22(3): 265–275 doi: 10.1006/cres.2001.0258 |
| Ghanmi, M., Ben Youssef, M., Jouirou, M., et al., 2001. Halocinèse Crétacée au Jebel Kebbouch (Nord-Ouest Tunisien): Mise en Place à Fleur d'eau et Evolution d'un «Glacier de sel» Albien, Comparaisons. Eclog. Geol. Helv. , 94: 153–160 http://www.researchgate.net/publication/285767838_Halocinese_cretacee_au_Jebel_Kebbouch_Nord-Ouest_tunisien_Mise_en_place_a_fleur_d'eau_et_evolution_d'un_glacier_de_sel_albien_comparaisons |
| Gorican, S., Smuc, A., Baumgartner, P. O., 2003. Toarcian Radiolaria from Mt. Mangart (Slovenian-Italian Border) and Their Paleoecological Implications. Marine Micropaleontology, 49(3): 275–301 http://www.sciencedirect.com/science/article/pii/S0377839803000343 |
| Guiraud, R., Maurin, J. C., 1992. Early Cretaceous Rifts of Western and Central Africa: An Overview. Tectonophysics, 213(1–2): 153–168 http://www.onacademic.com/detail/journal_1000035331809810_0e9e.html |
| Hammami, M., 1999. Tectonique, Halocinèse et Mise en Place de la Minéralisation dans la Zone des Diapirs (Tunisie Septentrionale): [Dissertation]. Université de Tunis II, Tunisie |
| Hancock, J., 2001. A Proposal for a New Position for the Aptian/Albian Boundary. Cretaceous Research, 22: 677–683 doi: 10.1006/cres.2001.0293 |
| Hardenbol, J., Thierry, J., Farley, M. B., et al., 1998. Mesozoic and Cenozoic Sequence Chronostratigraphic Framework of European Basins. Special Publication of Soc. Eco. Palaeont. Mineral, 60: 3–14 |
| Heldt, M., Lehmann, J., Willems, H., 2010. Calcareous Dinoflagellate Cysts from the Aptian/Albian Boundary Interval of Northern Germany: Abundance Patterns Related to Orbital Forcing? Newsletters on Stratigraphy, 44(1): 37–55 doi: 10.1127/0078-0421/2010/0003 |
| Herrle, J. O., Pross, J., Friedrich, O., et al., 2003. Forcing Mechanisms for Mid Cretaceous Black Shale Formation: Evidence from the Upper Aptian and Lower Albian of the Vocontian Basin (SE France). Palaeogeogr., Palaeoclimatol., Palaeoecol. , 190: 399–426 doi: 10.1016/S0031-0182(02)00616-8 |
| Holbourn, A. E. L., Kuhnt, W., Soeding, E., 2001. Atlantic Paeobathymetry, Paleoproductivity and Paleocirculation in the Late Albian: The Benthic Foraminiferal Record. Palaeogeogr., Palaeoclimatol., Palaeoecol. , 170: 171–196 doi: 10.1016/S0031-0182(01)00223-1 |
| Holbourn, A. E. L., Moullade, M., 1998. Lower Cretaceous Benthic Foraminifer Assemblages, Equatorial Atlantic: Biostratigraphic, Paleoenvironmental, and Paleobiogeographic Significance. In: Mascle, J., Lohmann, G. P., Moullade, M., eds., Proceedings of the Ocean Drilling Program, Scientific Results, 159: 347–362 |
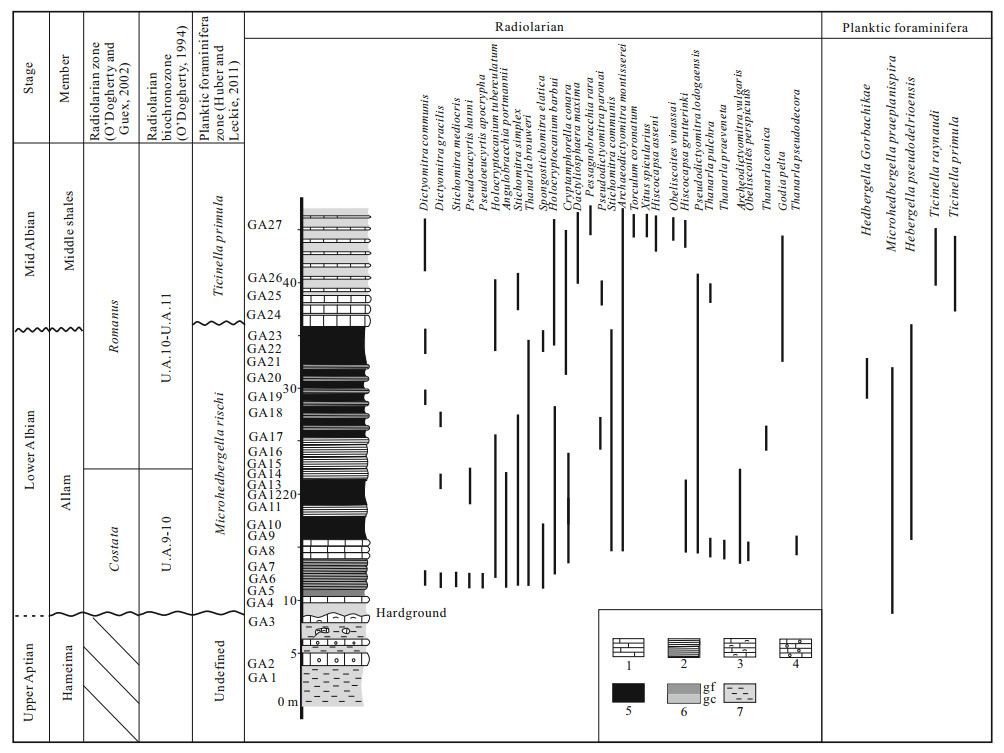
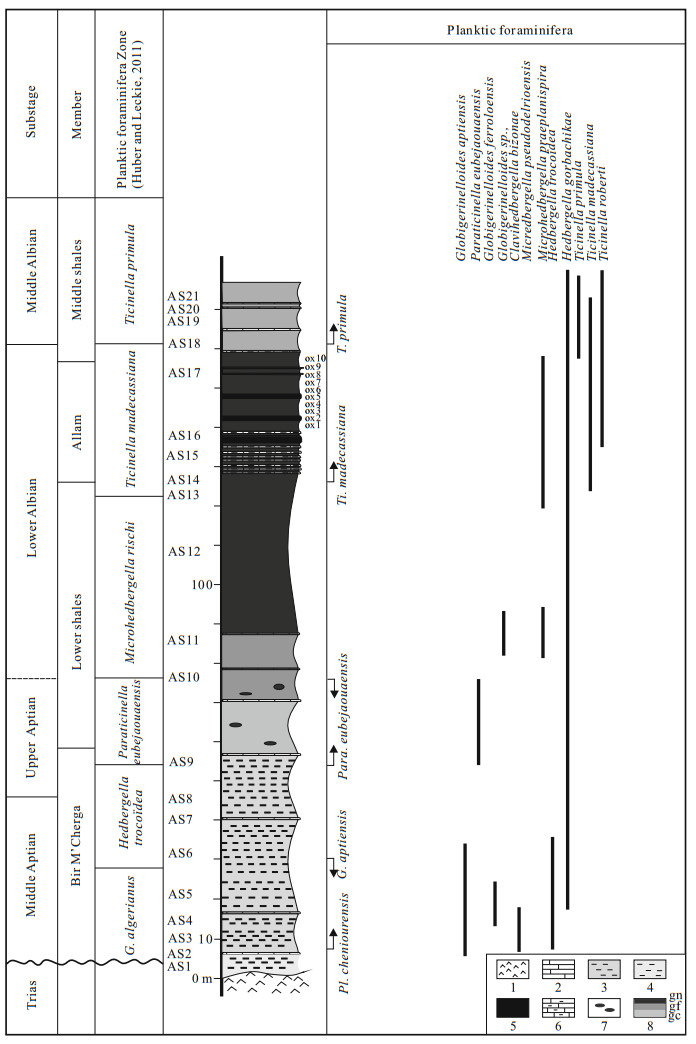
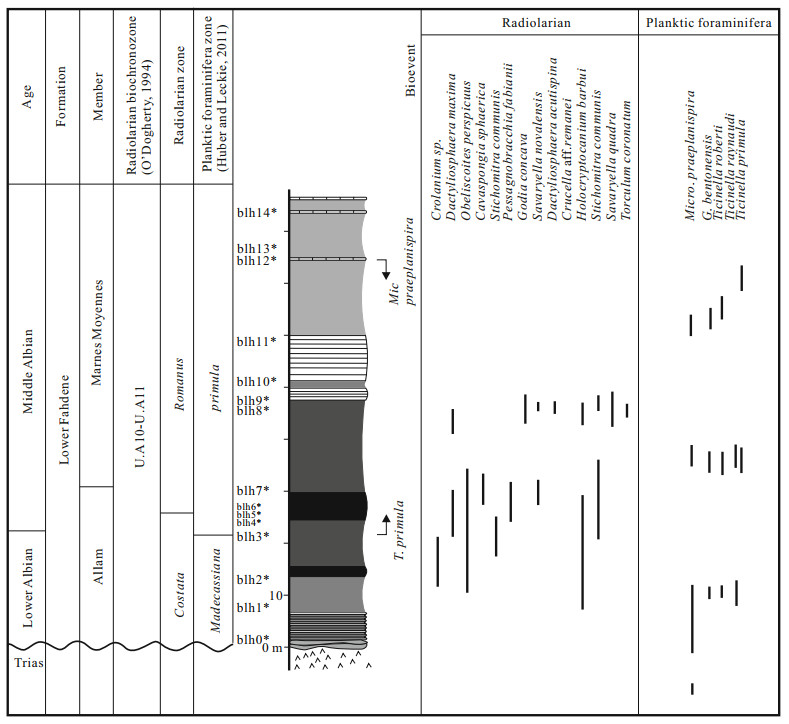
| Huber, B. T., Leckie, R. M., 2011. Planktic Foraminiferal Species Turnover across Deep-Sea Aptian/Albian Boundary Sections. J. Foram. Res. , 41(1): 53–95 doi: 10.2113/gsjfr.41.1.53 |
| Jauzein, A., 1967. Contribution à l'étude Géologique de la Tunisie Septentrionale: les Confins de la Dorsale Tunisienne. Ann. Min. Géol. , 22: 475 |
| Jud, R., 1994. Biochronology and Systematics of Early Cretaceous Radiolaria of the Western Tethys. Mem. Geol. (Lausanne), 19: 1–147 |
| Kariminia, S. M., 2006. Upper Jurassic and Lower Cretaceous Radiolaria Biostratigraphy of California Coast Ranges: [Dissertation]. University of Texas, Dallas |
| Kemkin, I. V., 2009. Early Cretaceous Radiolarians of the Amur River Lower Stream Area (Russia Far East) and Their Tectonic Significance. Acta Geosc. Sin. , 30(Suppl. 1): 21–24 http://www.cagsbulletin.com/dqxbcn/ch/reader/create_pdf.aspx?file_no=2009S114 |
| Kemkin, I. V., Filippov, A. N., 2010. Zyabrev's Critical Paper Entitled "On the Biostratigraphy of Accretionary Complexes of the Far East: A Critical Review of Several Papers. Russian Journal of Pacific Geology, 4(1): 91–94 doi: 10.1134/S1819714010010070 |
| Kennedy, W. J., Gale, A. S., Bown, P. R., et al., 2000. Integrated Stratigraphy across the Aptian-Albian Boundary in the Marnes Bleues, at the Col de Pré-Guittard, Arnayon (Drôme), and at Tartonne (Alpes de Haute-Provence), France: A Candidate Global Boundary Stratotype Section and Boundary Point for the Base of the Albian Stage. Cretaceous Research, 21: 591–720 doi: 10.1006/cres.2000.0223 |
| Khayati-Ammar, H., 1996. Stratigraphie et Micropaléontologie de la Série Crétacée Inférieur des Massifs Derdj-Bargou (Tunisie du Centre Nord). Mémoire DEA, Université de Tunis II, Tunisie |
| Khomsi, S., Bédir, M., Soussi, M., et al., 2004. Mise en Evidence et Analyse d'une Structure Atlasique Ennoyée au Front de la Chaîne Alpine Tunisienne. Comptes Rendus Geosci. , 336(14): 1293–1300 doi: 10.1016/j.crte.2004.05.003 |
| Kuhnt, H., Thurow, J., Wiedmann, J., et al., 1986. Oceanic Anoxic Conditions around the Cenomanian/Turonian Boundary and the Response of Biota. Mitteilungender Geologisch-Palaöntologisches Institut der Universität Hamburg, 60: 205–246 http://www.researchgate.net/publication/284668570_Oceanic_anoxic_conditions_around_the_CenomanianTuronian_boundary_and_the_response_of_the_biota |
| Kurilov, D. V., Vishnevskaya, V. S., 2011. Early Cretaceous Radiolarian Assemblages from the East Sakhalin Mountains. Stratigraphy and Geological Correlation, 19(1): 44–62 doi: 10.1134/S0869593810051028 |
| Lehmann, J., Heldt, M., Bachmann, M., et al., 2009. Aptian (Lower Cretaceous) Biostratigraphy and Cephalopods from North Central Tunisia. Cretaceous Research, 30(4): 895–910 doi: 10.1016/j.cretres.2009.02.002 |
| Meddeb, S., 1986. Sédimentationet Tectonique Polyphaséedans Lesdômes d'Enfidha Sahel Tunisien. Thèsededoctorat3e Cycle, Univ. Paris-Sud. 15 |
| Memmi, L., 1999. L'Aptien et l'Albien de Tunisie. Biostratigraphie à Partir des Ammonites. Bull. Soc. Geol. de France, 170(3): 303–309 |
| Michalík, J., Soták, J., Lintnerová, O., et al., 2008. The Stratigraphic and Paleoenvironmental Setting of Aptian OAE Black Shale Deposits in the Pieniny Klippen Belt, Slovak Western Carpathians. Cretaceous Research, 29(5-6): 871–892 doi: 10.1016/j.cretres.2008.05.005 |
| Moullade, M., Bellier, J. P., Tronchetti, G., 2002. Hierarchy of Criteria, Evolutionary Processes and Taxonomic Simplification in the Classification of Lower Cretaceous Planktonic Foraminifera. Cretaceous Research, 23(1): 111–148 doi: 10.1006/cres.2002.0304 |
| Musavu-Moussavou, B., Danelian, T., 2006. The Radiolarian Biotic Response to Oceanic Anoxic Event 2 in the Southern Part of the Northern Proto-Atlantic (Demerara Rise, ODP Leg 207). Rev. Micropal. , 49(3): 141–163 doi: 10.1016/j.revmic.2006.04.004 |
| Musavu-Moussavou, B., Danelian, T., Baudin, F., et al., 2007. The Radiolarian Biotic Response during OAE2. A High-Resolution Study across the Bonarelli Level at Bottaccione (Gubbio, Italy). Rev. Micropal. , 50: 253–287 |
| O'Dogherty, E. S., Carter, P., Dumitrica, S., et al., 2009. Catalogue of Mesozoic Radiolarian Genera. Part 2: Jurassic-Cretaceous. Geodiversitas, 31: 271–356 http://sciencepress.mnhn.fr/sites/default/files/articles/pdf/g2009n2a1.pdf |
| O'Dogherty, L., 1994. Biochronology and Paleoecology of Mid-Cretaceous Radiolarians from Northern Apennines (Italy) and Betic Cordillera (Spain). Mémoires de Géologie (Lausanne), 21: 415 http://www.unil.ch/files/live/sites/iste/files/shared/X.Library/Memoirs%20of%20Geology/21%20-%20ODogherty%20(1994).pdf |
| O'Dogherty, L., Bill, M., Gorican, S., et al., 2006. Bathonian Radiolarians from an Ophiolitic Mélange of the Alpine Tethys (Gets Nappe, Swiss-French Alps). Micropaleontology, 51(6): 425–485 http://www.onacademic.com/detail/journal_1000036371966810_5f0c.html |
| O'Dogherty, L., Carter, E. S., Gorican, S., et al., 2010. Triassic Radiolarian Biostratigraphy. Geological Society, London, Special Publications, 334: 163–200 doi: 10.1144/SP334.8 |
| O'Dogherty, L., Guex, J., 2002. Rates and Pattern of Evolution among Cretaceous Radiolarians: Relations with Global Paleoceanographic Events. Micropaleontology, 48: 1–22 doi: 10.2113/48.1.1 |
| Palechek, T. N., Savel'ev, D. P., Savel'eva, O. L., 2010. Albian-Cenomanian Radiolarian Assemblage from the Smaginsk Formation, the Kamchatskii Mys Peninsula of Eastern Kamchatka. Stratigraphy and Geological Correlation, 18(1): 63–82 doi: 10.1134/S0869593810010053 |
| Perthuisot, V., 1978. Dynamique et Pétrogenèse des Extrusions Triasiques en Tunisie Septentrionale: [Dissertation]. Ecole Normale Supérieure, Paris |
| Reichelt, K., 2005. Late Aptian-Albian of the Vocontian Basin (SE-France) and Albian of NE-Texas: Biostratigraphic and Paleoceanographic Implications by Plankticforaminifera Faunas: [Dissertation]. Erlangung des Grades eines Doktors der Naturwissenschaften, Université de Tübingen. 125 |
| Rigane, A., Feki, M., Gourmelen, C., et al., 2010. The «Aptian Crisis» of the South-Tethyan Margin: New Tectonic Data in Tunisia. Journal of African Earth Sciences, 57(4): 360–366 doi: 10.1016/j.jafrearsci.2009.11.005 |
| Robaszynski, F., Amedro, F., Gonzalez-Donoso, J. M., et al., 2008. The Albian (Vraconnian)-Cenomanian Boundary at the Western Tethyan Margins (Central Tunisia and Southeastern France). Bull. Soc. Géol. Fr. , 179(3): 245–256 doi: 10.2113/gssgfbull.179.3.245 |
| Robaszynski, F., Caron, M., Amédro, F., et al., 1994. Le Cénomanien de la Région de Kalaat Senan (Tunisie Centrale): Litho-Biostratigraphie et Interpré-Tation Séquentielle. Revue de Paléobiologie, 12(2): 351–505 |
| Saadi, J. M., Ben Youssef, P., Souquet, B., et al., 1994. Stratigraphie Séquentielle du Crétacé Inférieur de la Région d'Enfidha (NE de la Tunisie). Comp. Ren. Acad. Sci. de Paris, 319: 119–125 |
| Saadi, J., 1990. Exemple de Sédimentation Syntectonique au Crétacé Inférieur le Long d'une Zone de Décrochement NS. Les Structures d'Enfidha (Tunisie Nord-Orientale). Géodynamique, 5: 17–33 |
| Saadi, J., 1991. Sedimentation en Zone Mobile Coulissante, l'exemple du Crétacé Inferieur des Structures Submeridiennes de La région d'Enfidha. (Prolongement Septentrional de l'Axe N. S-Tunisie Nord-Orientale): [Dissertation]. Université de Pau, France |
| Saadi, J., Duee, G., 1991. Importance des Phenomenes de Resedimentation au Passage Jurassique-Crétace et Aptien-Albien dans les Structures d'Enfidha (Tunisie Nord-Orientale) and Consequences Tectono-Eustatiques. Geol. Medit. , 18: 135–147 http://www.researchgate.net/publication/327724513_Importance_des_phenomenes_de_resedimentation_au_passage_Jurassique-Cretace_et_Aptien-Albien_dans_les_structures_d'Enfidha_Tunisie_nord-orientale_Consequences_tectono-eustatiques |
| Schlanger, S. O., Jenkyns, H. C., 1976. Cretaceous Oceanic Anoxic Events: Causes and Consequences. Geol. Mijnb. , 55(3–4): 179–184 http://ci.nii.ac.jp/naid/10011370228 |
| Silva, I. P, Sliter, W. V., 1999. Cretaceous Paleoceanography: Evidence from Planktic Foraminiferal Evolution. In: Barrera, E., Johnson, C. C., eds., Evolution of the Cretaceous Ocean-Climate System. Geol. Soc. Am., Special Paper, 332: 301–328 |
| Slaczka, A., Gasiñski, M. A., Bąk, M., et al., 2009. The Clasts of Cretaceous Marls in the Conglomerates of the Konradsheim Formation (Pöchlau Quarry, Gresten Klippen Zone, Austria). Geol. Carpath. , 60: 151–164 doi: 10.2478/v10096-009-0010-7 |
| Sliter, W. V., 1989. Biostratigraphic Zonation for Cretaceous Planktic Foraminifers Examined in Thin Section. J. Foram. Res. , 19(1): 1–19 doi: 10.2113/gsjfr.19.1.1 |
| Soua, M., Zaghbib-Turki, D., O'Dogherty, L., 2006. Les Réponses Biotiques des Radiolaires à l'événement Anoxique du Cénomanien Supérieur dans la Marge sud Téthysienne (Tunisie). Proceeding of the 10th Tunisian Petroleum Exploration & Production Conference. 195–216 |
| Talbi, R., 1991. Etude Géologique et Géochimique des Faciès Riches en Matière Organique d'âge Albien du Bassin de Bir M'Cherga (NE de Tunisie): Déterminisme de leur Genèse et Intérêt pétrolier de la Région: [Dissertation]. University de Tunis, Tunisie |
| Thurow, J., 1988. Cretaceous Radiolarians of the North Atlantic Ocean: ODP Leg 103 (Sites 638, 640 and 641) and DSDP Legs 93 (Site 603) and 47B (Site 398). In: Boillot, G., Wintere, E. L., et al., eds., Proceedings of the Ocean Drilling Program, Scientific Results, 103: 379–418 |
| Tsikos, H., Karakitsios, V., van Breugel, Y., et al., 2004. Organic Carbon Deposition in the Cretaceous of the Ionian Basin, NW Greece: The Paquier Event (OAE 1b) Revisited. Geological Magazine, 141(4): 401–416 doi: 10.1017/S0016756804009409 |
| Tyszka, J., 2006. Taxonomy of Albian Gavelinellidae (Foraminifera) from the Lower Saxony Basin, Germany. Palaeontology, 49(6): 1303–1334 doi: 10.1111/j.1475-4983.2006.00602.x |
| Vila, J. M., Ben Youssef, M., Charrière, A., et al., 1994. Découverte en Tunisie, au SW du Kef, de Matériel Triasique Interstratifié dans l'Albien: Extension du Domaine à" Glaciers de sel" Sous-Marins des Confins Algéro-Tunisiens. Comp. Ren. Acad. Sci. , 318(II): 1661–1667 |
| Vila, J. M., Ben Youssef, M., Chikhaoui, M., et al., 1996. Deuxième étude de Surface d'un Grand «Glacier de Sel» Sous-Marin Albien (250 km2): les Masses Triasiques du «Diapir» de Ben Gasseur et de l'anticlinal du Kef (NW Tunisien). Bull Soc. Géol. Fr. , 167: 235–246 http://cat.inist.fr/?aModele=afficheN&cpsidt=2505908 |
| Vila, J. M., Ghanmi, M., Ben Youssef, M., et al., 2002. Les «Glaciers de Sel» Sous-Marins des Marges Continentales Passives du Nord-Est du Maghreb (Algérie-Tunisie) et de la Gulf Coast (USA): Comparaisons, Nouveau Regard sur les «Glaciers de Sel Composites, Illustré par celui de Fedj el Adoum (Nord-Ouest Tunisien) et Revue Globale». Eclogae Geol. Hel. , 95: 347–380 |
| Yurtsever, T. S., Tekin, U. K., Demirel, I. H., 2003. First Evidence of the Cenomanian/Turonian Boundary Event (CTBE) in the Alakircay Nappe of Antalya Nappes, Southwest Turkey. Cretaceous Research, 24: 41–53 doi: 10.1016/S0195-6671(03)00021-1 |
| Zghal, I., Ben Hadjali, N., Razgallah, S., et al., 1997. Foraminifères et Ostracodes de l'Aptien-Albien du Jebel Hameima (Region de Tadjerouine, Tunisie): Biostratigraphie, Paleoécologie. Africa Geosc. Rev. , 4: 361–372 |
| Zyabrev, S. V., 2011. Stratigraphy and Structure of the Central East Sakhalin Accretionary Wedge (Eastern Russia). Russian Journal of Pacific Geology, 5(4): 313–335 doi: 10.1134/S1819714011040087 |
| Zyabrev, S. V., Aitchison, J. C., Abrajevitch, A. V., et al., 2003. Precise Radiolarian Age Constraints on the Timing of Ophiolite Generation and Sedimentation in the Dazhuqu Terrane, Yarlung-Tsangpo Suture Zone, Tibet. J. Geol. Soc. London, 160(6): 591–599 http://ci.nii.ac.jp/naid/20000877865 |
Figures(6)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: