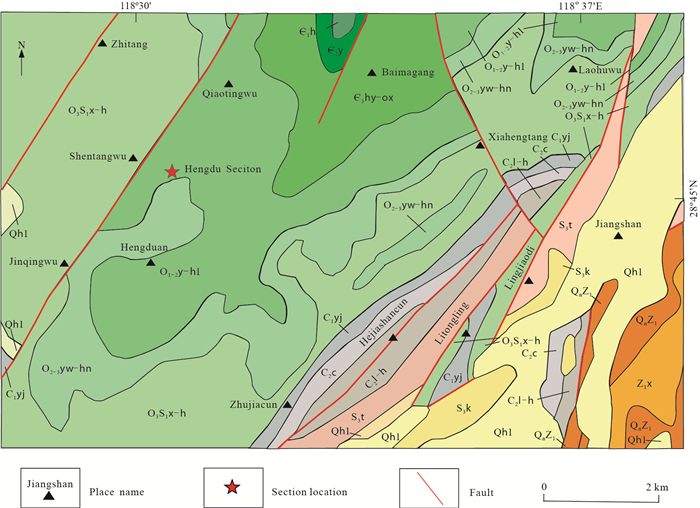
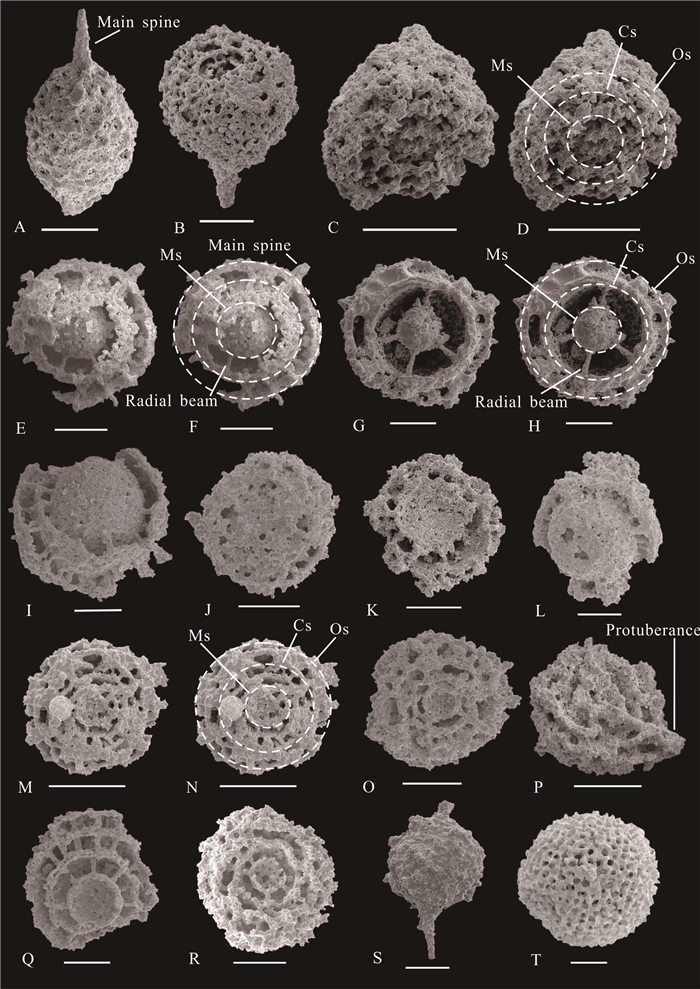
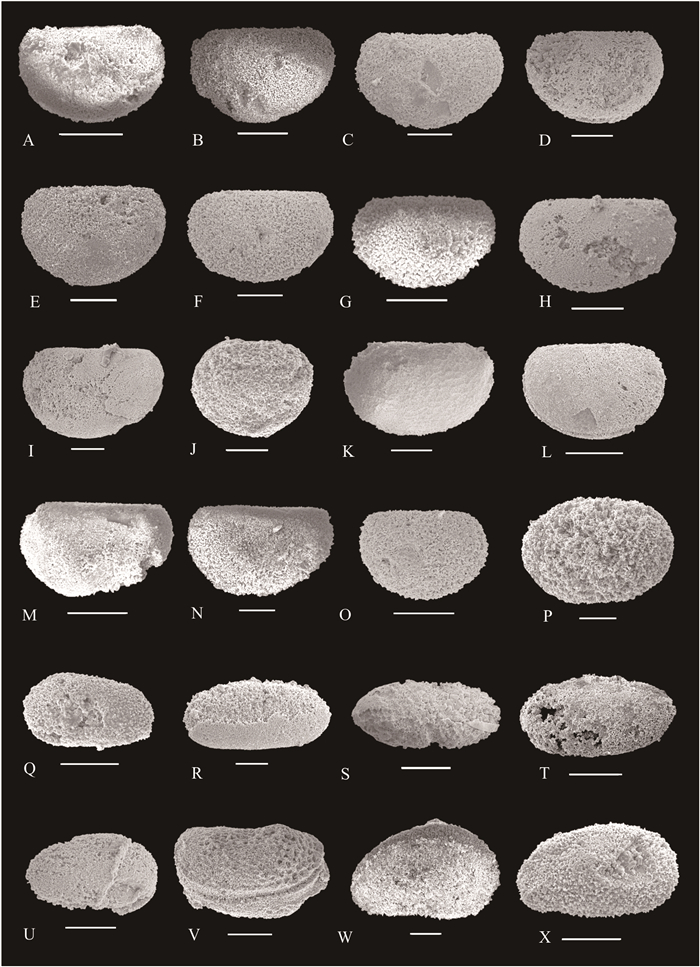
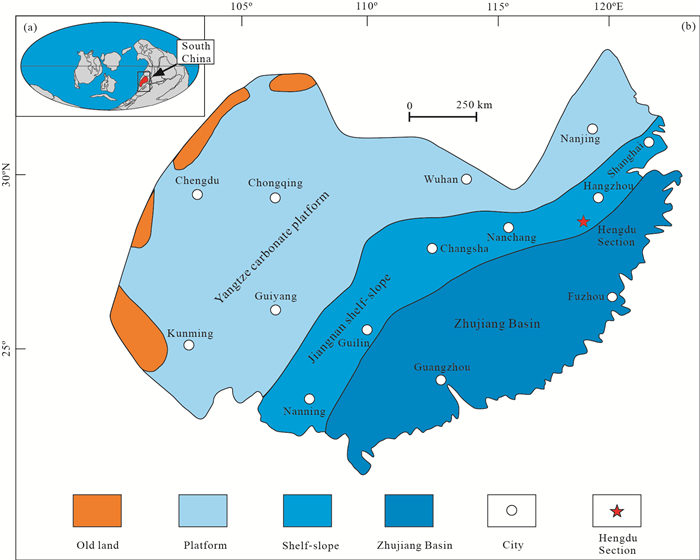
| Citation: | Yuhao Yi, Aihua Yuan, Jonathan C Aitchison, Qinglai Feng. Upper Darriwilian (Middle Ordovician) Radiolarians and Ostracods from the Hulo Formation, Zhejiang Provicnce, South China. Journal of Earth Science, 2018, 29(4): 886-899. doi: 10.1007/s12583-017-0951-6

|

| Aitchison, J. C., Flood, P. G., Malpas, J., 1998. Lowermost Ordovician (Basal Tremadoc) Radiolarians from the Little Port Complex, Western Newfoundland. Geological Magazine, 135(3): 413-419. doi: 10.1017/s001675689800867x |
| Bandel, K., Becker, G., 1975. Ostracoden aus Paläozoischen-Pelagischen Kalken der Karnischen Alpen (Silurium bis Unterkarbon). Senkenbergiana Lethaea, 56(1): 1-83 (in German) doi: 10.1007/BF03042773.pdf |
| Berdan, J. M., 1976. Middle Ordovician Leperditicopid Ostracodes from the Ibex Area, Millard County, Western Utah.Brigham Young University College Studies, 23: 37-65 doi: 10.2307/4541260 |
| Berdan, J. M., 1984. Leperditicopid Ostracodes from Ordovician Rocks of Kentucky and Nearby States and Characteristic Features of the Order Leperditicopida. United States Government Printing Office, Washington DC. 1-40 |
| Berdan, J. M., 1988. Middle Ordovician (Whiterockian) Palaeocopid and Podocopid Ostracodes from the Ibex Area, Millard County, Western Utah. New Mexico Bureau of Mines and Mineral Resources Memoir, 44: 273-301 doi: 10.2307/4541260 |
| Braddy, S. J., Tollerton, V. P., Racheboeuf, P. R., et al., 2004. Eurypterids, Phyllocarids, and Ostracodes. In: Webby, B. D., Paris, F., Droser, M. L., et al., eds., The Great Ordovician Biodiversification Event. Columbia University Press, New York. 255-256. doi: 10.7312/webb12678-026 |
| Buckman, S., Aitchison, J. C., 2001. Middle Ordovician (Llandeilan) Radiolarians from West Junggar, Xinjiang, China. Micropaleontology, 47(4): 359-367. doi: 10.2113/47.4.359 |
| Cui, Z. L., Huan, H., Song, Q. Y., 2000. The Late Ordovician Radiolarian Assemblage of the North Qinling Back-Arc Basin, China. Acta Geologica Sinica, 74(3): 254-258 (in Chinese with English Abstract) https://www.researchgate.net/publication/240670968_Magmagenesis_in_the_Ordovician_backarc_basins_of_the_Northern_Qilian_Mountains_China |
| Danelian, T., 1999. Taxonomic Study of Ordovican (Llanvirn-Caradoc) Radiolaria from the Southern Uplands (Scotland, U.K.). Geodiversitas, 21(4): 625-635 |
| Danelian, T., Clarkson, E. N. K., 1998. Ordovician Radiolaria from Bedded Cherts of the Southern Uplands. Scottish Journal of Geology, 34(2): 133-137. doi: 10.1144/sjg34020133 |
| Danelian, T., Floyd, J. D., 2001. Progress in Describing Ordovician Siliceous Biodiversity from the Southern Uplands (Scotland, U.K.). Transactions of the Royal Society of Edinburgh: Earth Sciences, 91(3/4): 489-498. doi: 10.1017/s0263593300008336 |
| Danelian, T., Noble, P., Pouille, L., et al., 2013. Palaeogeographical Distribution of Ordovician Radiolarian Occurrences: Patterns, Significance and Limitations. Geological Society, London, Memoirs, 38(1): 407-413. doi: 10.1144/m38.25 |
| Danelian, T., Popov, L. E., 2003. Ordovician Radiolarian Biodiversity: Insights Based on New and Revised Data from Kazakhstan. Bulletin de la Société Géologique de France, 174(4): 325-335. doi: 10.2113/174.4.325 |
| Danelian, T., Popov, L. E., Tolmacheva, T. Y., et al., 2011. Ordovician Radiolaria and Conodonts from the Peri-Gondwanan Karatau-Naryn Microplate (Sarydzhaz, Eastern Kyrgyzstan). Geobios, 44(6): 587-599. doi: 10.1016/j.geobios.2011.02.006 |
| Fang, Z. J., 2006. An Introduction to Ordovician Bivalves of Southern China, with a Discussion of the Early Evolution of the Bivalvia. Geological Journal, 41(3/4): 303-328. doi: 10.1002/gj.1048 |
| Feng, Z. Z., Peng, Y. M., Jin, Z. K., et al., 2001. Lithofacies Palaeogeography of the Middle and Late Ordovician in South China. Journal of Palaeogeography, 3(4): 10-24 (in Chinese with English Abstract). http://www.sciencedirect.com/science/article/pii/S2095383615301188 |
| Feng, Z. Z., Peng, Y. M., Jin, Z. K., et al., 2003. Lithofacies Palaeogeography of the Middle Ordovician China. Journal of Palaeogeography, 5(3): 263-278 (in Chinese with English Abstract) https://www.researchgate.net/publication/282541854_The_Lithofacies_Palaeogeography_and_Control_of_the_Reservoirs_in_Middle_Ordovician_Ma541_Submember_in_Western_Jingbian_Tableland |
| Frank, M. S., 1969. Ordovician Ostacode Aparchites Whiteavesi Jones (1889), and Problems of Relationships. Journal of Paleontology, 43(5): 1237-1244 |
| Ghobadi Pour, M., Williams, M., Vannier, J., et al., 2006. Ordovician Ostracods from East Central Iran. Acta Palaeontologica Polonica, 51(3): 551-560 http://yadda.icm.edu.pl/yadda/element/bwmeta1.element.dl-catalog-b6c39f22-2860-476c-8d3f-cc9b1c30c7ab |
| Ghobadi Pour, M., Mohibullah, M., Williams, M., et al., 2011. New, Early Ostracods from the Ordovician (Tremadocian) of Iran: Systematic, Biogeographical and Palaeoecological Significance. Alcheringa: An Australasian Journal of Palaeontology, 35(4): 517-529. doi: 10.1080/03115518.2011.538909 |
| Goto, H., Ishiga, H., 1991. Study of Late Ordovician Radiolarians from the Lachlan Fold Belt, Southeastern Australia. Geological Reports, Shiname University, 10: 57-62 |
| Goto, H., Umeda, M., Ishiga, H., 1992. Late Ordovician Radiolarians from the Lachlan Fold Belt, Southeastern Australia. Memoirs of the Faculty of Science, Shimane University, 26: 145-170 |
| Guo, F. S., 1994. On Collating and Applying of the Lithostratigraphic Units in Jiangshan Area, Zhejiang. Journal of East China Geological Institute, 17(3): 254-263 (in Chinese with English Abstract) |
| Guo, F. S., 1998. The Ordovician Sedimentary Facies and Palaeogeography and Their Tectonic Controls in Jiangshan, Zhejiang. Sedimentary and Palaeogeography, 18(4): 57-62 (in Chinese with English Abstract) https://www.researchgate.net/profile/Wenkun_Qie |
| Henningsmoen, G., 1953. Classification of Paleozoic Straight-Hinge Ostracods.Norske Geologische Tidsskrift, 31: 185-288 https://deepblue.lib.umich.edu/bitstream/handle/2027.42/48257/ID096.pdf;sequence=2 |
| Hessland, I., 1949. Lower Ordovician Ostracodes of the Siljan District, Sweden. Bulletin of the Geological Institution of the University of Uppsala, 33: 408 https://www.researchgate.net/publication/267638645_Terminal_Ordovician_stratigraphy_of_the_Siljan_district_Sweden |
| Hou, Y. T., 1956a. Some New Species of Ostracods from Middle Ordovician. Acta Palaeontologica Sinica, 4(3): 355-360 (in Chinese with English Abstract) |
| Hou, Y. T., 1956b. Ordovician Ostracoda from Western Zhejiang. Acta Palaeontologica Sinica, 4(4): 535-597 (in Chinese with English Abstract) https://www.researchgate.net/publication/233098729_Early_Sandbian_Late_Ordovician_conodonts_from_the_Yenwashan_Formation_western_Zhejiang_South_China |
| Hu, K. M., Tang, Z. C., Meng, X. S., et al., 2016. Chronology of Petrogenesis and Mineralization of Datongkeng Porphyry W-Mo Deposit in West Zhejiang. Earth Science, 41(9): 1435-1450. doi:103799/dqkx2016502 (in Chinese with English Abstract) |
| Iwata, K., Schmidt, B. L., Leitch, E. C., et al., 1995. Ordovidan Microfossils from the Ballast Formation (Girilambone Group) of New South Wales. Australian Journal of Earth Sciences, 42(4): 371-376. doi: 10.1080/08120099508728208 |
| Jannusson, V., 1957. Middle Ordovician Ostracodes of Central and Southern Sweden. Bulletin Geological Institution of the University of Uppsala, 37: 173-442 https://www.researchgate.net/publication/233369763_Trilobites_and_stratigraphy_of_the_Middle_Ordovician_Killerod_Formation_Scania_Sweden |
| Kempf, E. K., 2009. Paraparchites Eskridgensis, New Name for the Ostracode Paraparchites Punctatus Watabe and Kaesler, 2004. Journal of Paleontology, 83(3): 501. doi: 10.1666/09-005.1 |
| Kesling, R. V., Crafts, F. S., Darby, D. G., et al., 1960. Middle Ordovician Black River Ostracods from Michigan, Introduction and Part Ⅰ. Museum of Paleontology the University of Michigan, XV(13): 293-314 https://deepblue.lib.umich.edu/bitstream/handle/2027.42/48343/ID184.pdf;sequence=2 |
| Kesling, R. V., 1960. Middle Ordovician Black River Ostracods from Michigan, Part Ⅱ. Museum of Paleontology the University of Michigan, XV(15): 349-363 https://www.researchgate.net/publication/30849103_Middle_Ordovician_Black_River_Ostracods_from_Michigan_Part_III_Platybolbina |
| Knüpfer, J., 1968. Ostracoden aus dem Oberen Ordovizium Thüringens. Freiberger Forschung Hefte, C234: 5-29 (in German) |
| Kozur, H., 1972. Die Bedeutung Triassischer Ostracoden für Stratigraphische und Paläökologische Untersuchungen.Mitteiungen Gesellschaft Geologie Bergbaustelle, 21: 623-660 (in German) |
| Kuwano, Y., 1987. Early Devonian Conodonts and Ostracodes from Central Japan. Bulletin of Natural. Science Mususem., Tokyo, 13(2): 77-105 http://jpaleontol.geoscienceworld.org/content/64/4/595.abstract |
| Lai, C. G., Jin, R. G., Lin, B. Y., et al., 1993. Biofacies, Sedimentary Facies and Palaeogeographic Characteristics of the Ordovician in the Lower Yangtze Area. Geological Publishing Press, Beijing. 1-84 (in Chinese) |
| Landing, E., Mohibullah, M., Williams, M., 2013. First Middle Ordovician Ostracods from Western Avalonia: Paleogeographical and Paleoenvironmental Significance. Journal of Paleontology, 87(2): 269-276. doi: 10.1666/12-065r1.1 |
| Levinson, S. A., 1961. New Genera and Species of Bromide (Middle Ordovician) Ostracodes of Oklahoma. Micropaleontology, 7(3): 359-364. doi: 10.2307/1484369 |
| Li, H., 1995. New Genara and Species of Middle Ordovician Nassellaria and Albaillellaria from Baijingsi, Quilian Mountains, China. Scientia Geologica Sinica, 4(3): 331-346 |
| Li, J., Servais, T., Yan, K., et al., 2007. Microphytoplankton Diversity Curves of the Chinese Ordovician. Bulletin de la Societe Geologique de France, 178(5): 399-409. doi: 10.2113/gssgfbull.178.5.399 |
| Liu, C. G., Li, G. R., Wang, D. W., et al., 2016. Middle-Upper Ordovician (Darriwilian-Early Katian) Positive Carbon Isotope Excursions in the Northern Tarim Basin, Northwest China: Implications for Stratigraphic Correlation and Paleoclimate. Journal of Earth Science, 27(2): 317-328. doi: 10.1007/s12583-016-0696-2 |
| Liu, W., Xu, X. S., Feng, X. T., et al., 2010. Radiolarian Siliceous Rocks and Palaoenvironmental Reconstruction for the Upper Ordovician Wufeng Formation in the Middle-Upper Yangtze Area. Sedimentary Geology and Tethyan Geology, 30(3): 65-70 (in Chinese with English Abstract) https://www.researchgate.net/scientific-contributions/2024236145_Dazhong_Dong |
| Lu, Y., Ma, Y., Wang, Y., et al., 2017. The Sedimentary Response to the Major Geological Events and Lithofacies Characteristics of Wufeng Formation-Longmaxi Formation in the Upper Yangtze Area. Earth Science, 42(7): 1169-1184. doi:10/3799/dqkx.2017.095 (in Chinese with English Abstract) |
| Luo, S., Gao, Z., He, Y., et al., 2002. Ordovician Carbonate Contourite Drifts in Hunan and Gansu Provinces, China. In: Stow, D. A. V., ed., Deep-Water Contourite Systems: Modern Drifts and Ancient Series, Seismic and Sedimentary Characteristics. Geological Society Memoirs, London, 22(1): 433-442. doi: 10.1144/gsl.mem.2002.022.01.30 |
| Maletz, J., 2007. The Early Ordovician Beothuka Terranova (Radiolaria) Faunal Assemblage in Western Newfoundland. Paläontologische Zeitschrift, 81(1): 71-82. doi: 10.1007/bf02988380 |
| Maletz, J., 2011. Radiolarian Skeletal Structures and Biostratigraphy in the Early Palaeozoic (Cambrian-Ordovician). Palaeoworld, 20(2/3): 116-133. doi: 10.1016/j.palwor.2010.12.007 |
| Maletz, J., Albanesi, G. L., Voldman, G. G., 2009. Lower Darriwilian Radiolarians from the Argentine Precordillera. Geobios, 42(1): 53-61. doi: 10.1016/j.geobios.2008.09.002 |
| Maletz, J., Bruton, D. L., 2005. The Beothuka Terranova (Radiolaria) Assemblage and Its Importance for the Understanding of Early Ordovician Radiolarian Evolution. Geological Magazine, 142(6): 711-721. doi: 10.1017/s0016756805001391 |
| Maletz, J., Bruton, D. L., 2007. Lower Ordovician (Chewtonian to Castlemainian) Radiolarians of Spitsbergen. Journal of Systematic Palaeontology, 5(3): 245-288. doi: 10.1017/s1477201907002039 |
| Maletz, J., Bruton, D. L., 2008. The Middle Ordovician Proventocitum Procerulum, Radiolarian Assemblage of Spitsbergen and Its Biostratigraphic Correlation. Palaeontology, 51(5): 1181-1200. doi: 10.1111/j.1475-4983.2008.00803.x |
| Meidla, T., 1996a. Late Ordovician Ostracodes of Estonia. Fossilia Baltica, 2: 1-222 |
| Meidla, T., 1996b. Latest Ordovician Ostracodes of Baltoscandia. Geological Survey of Denmark and Greenland, 20: 65-71 doi: 10.1080/11035898109455206 |
| Meidla, T., 2007. Ostracods from the Upper Ordovician Borenshult Fauna, Sweden. GFF, 129(2): 123-132. doi: 10.1080/11035890701292123 |
| Melnikova, L. M., 1999. Ostracodes from the Billingen Horizon (Lower Ordovician) of the Leningrad Region. Paleontological Journal, 33: 147-152 https://www.researchgate.net/publication/289416648_The_lower_ordovician_Volkhov_and_Kunda_horizons_and_characteristics_of_trilobites_and_ostracodes_the_Volkhov_River_Leningrad_region |
| Melnikova, L. M., 2010. Some Ostracodes from the Gur'yanovka Formation (Upper Ordovician) of Northeastern Gorny Altai. Paleontological Journal, 44(4): 399-408. doi: 10.1134/s0031030110040064 |
| Melnikova, L. M., Tolmacheva, T. Y., Ushatinskaya, G. T., 2010. Find of Tremadocian Ostracodes in Cherts of Kazakhstan. Paleontological Journal, 44(1): 36-40. doi: 10.1134/s0031030110010053 |
| Mohibullah, M., Vandenbroucke, T. R. A., Williams, M., et al., 2011. Late Ordovician (Sandbian) Ostracods from the Ardwell Farm Formation, SW Scotland. Scottish Journal of Geology, 47(1): 57-66. doi: 10.1144/0036-9276/01-428 |
| Molostovskaya, I. I., 2010. Ostracodes from the Upper Permian Khivach Formation in Kolyma-Omolon Basin. Paleontological Journal, 44(3): 282-286. doi: 10.1134/s0031030110030068 |
| Nazarov, B. B., Popov, L. Y., 1976. Radiolarians, Inarticulate Brachiopods and Organisms of Uncertain Systematic Positon from the Middle Ordovician of Eastern Kazakhstan. Paleontologicheskiy Zhurnal, 4(4): 33-42 |
| Nazarov, B. B., Popov, L. Y., Apollonov, M. K., 1977. Lower Paleozoic Radiolarians of Kazakhstan. International Geology Review, 19(8): 913-920. doi: 10.1080/00206817709471089 |
| Nazarov, B. B., Popov, L. Y., 1980. Stratigraphy and Fauna of the Siliceous-Carbonate Sequence of the Ordovician of Kazakhstan (Radiolarians and Inarticulate Brachiopods). Trudy Geologicheskiy Institut Akademiy a Nauk SSR, 331: 1-190 (in Russian) http://www.academia.edu/2553858/Middle_Ordovician_Llandeilan_radiolarians_from_West_Junggar_Xinjiang_China |
| Nazarov, B. B., Ormiston, A. R., 1986. Trends in the Development of Paleozoic Radiolaria. Marine Micropaleontology, 11(1/2/3): 3-32. doi: 10.1016/0377-8398(86)90003-4 |
| Noble, P., Danelian, T., 2004. Ordovician Radiolarian Biodiversity Estimates. In: Webby, B. D., Paris, F., Droser, M. L., et al., eds., The Great Ordovician Biodiversification Event. Columbia University Press, New York. 97-101 |
| Noble, P. J., Webby, B. D., 2009. Katian (Ordovician) Radiolarians from the Malongulli Formation, New South Wales, Australia: A Reexamination. Journal of Paleontology, 83(4): 548-561. doi: 10.1666/08-179r.1 |
| Olempska, E., 1994. Ostracods of the Mójcza Limestone. Palaeontologia Polonica, 53: 129-212 |
| Öpik, A. A, 1935. Ostracoda from the Lower Ordovician Megalaspis- Limestone of Estonia and Russia. Tart ülikooli j.o. Loodusuurujate Seltsi Aruanded, Annales Societatis Rebus Naturae Investigandis in Universitate Tartu Constitutae, 42: 28-38 (in Russian) http://www.academia.edu/19082051/Phylogenetic_relationships_of_Early-Middle_Ordovician_ostracods_of_Baltoscandia |
| Peng, Y., Peng, Y. B., Lang, X. G., et al., 2016. Marine Carbon-Sulfur Biogeochemical Cycles during the Steptoean Positive Carbon Isotope Excursion (SPICE) in the Jiangnan Basin, South China. Journal of Earth Science, 27(2): 242-254. doi: 10.1007/s12583-016-0694-4 |
| Pouille, L., Danelian, T., Popov, L. E., 2014. A Diverse Upper Darriwilian Radiolarian Assemblage from the Shundy Formation of Kazakhstan: Insights into Late Middle Ordovician Radiolarian Biodiversity. Journal of Micropalaeontology, 33(2): 149-163. doi: 10.1144/jmpaleo2014-008 |
| Reynolds, L., 1987. The Taxonomy and Palaeoecology of Ostracodes from the Devonian Receptaculites Limestone, Taemas, New South Wales, Australia. Palaeontographica A, 162(3-6): 144-203 |
| Salas, M. J., Vannier, J., Williams, M., 2007. Early Ordovician Ostracods from Argentina: Their Bearing on the Origin of Binodicope and Palaeocope Clades. Journal of Paleontology, 81(6): 1384-1395. doi: 10.1666/05-134.1 |
| Salas, M. J., 2011. Early Ordovician (Floian) Ostracods from the Cordillera Oriental, Northwest Argentina. Geological Journal, 46(6): 637-650. doi: 10.1002/gj.1319 |
| Salas, M. J., Vaccari, N. E., 2012. New Insights into the Early Diversification of the Ostracoda: Tremadocian Ostracods from the Cordillera Oriental, Argentina. Acta Palaeontologica Polonica, 57(1): 175-190. doi: 10.4202/app.2009.1110 |
| Schallreuter, R., Suttner, T. J., Hinz-Schallreuter, I., 2005. Late Ordovician Ostracodes from Himalaya and Their Palaeobiogeographic Relations. Berliner Paläobiologische Abhandlungen, 6: 107-108 http://core.ac.uk/display/9672277 |
| Schallreuter, R., Hinz-Schallreuter, I., Suttner, T., 2008. New Ordovician Ostracodes from Himalaya and Their Palaeobiological and Palaeogeographical Implications. Revue de Micropaléontologie, 51(3): 191-204. doi: 10.1016/j.revmic.2007.02.003 |
| Schallreuter, R., Hinz-Schalleuter, I., 2009. Ostracod as a Tool for Palaeogeographic Reconstruction in the Ordovician. Institution of Erdwissensch, Berlin. 14 |
| Scotese, C. R., 2001. Atlas of Earth History, Volume 1, Paleogeography, PALEOMAP Project. University of Texas, Arlington. 52 |
| Scott, H. W., 1959. Type Species of Paraparchites Ulrich & Bassler. Journal of Paleontology, 33(4): 679-674 http://pubs.er.usgs.gov/pubs/pp/pp711B |
| Servais, T., Lehnert, O., Li, J., et al., 2008. The Ordovician Biodiversification: Revolution in the Oceanic Trophic Chain. Lethaia, 41(2): 99-109. doi: 10.1111/j.1502-3931.2008.00115.x |
| Servais, T., Owen, A. W., Harper, D. A. T., et al., 2010. The Great Ordovician Biodiversification Event (GOBE): The Palaeoecological Dimension. Palaeogeography, Palaeoclimatology, Palaeoecology, 294(3/4): 99-119. doi: 10.1016/j.palaeo.2010.05.031 |
| Shi, C. G., Wang, D. H., 1985. Middle Ordovician Ostracodes from Huanxian, Gansu. Northwest Geoscience, 10: 96-104 (in Chinese with English Abstract) https://www.researchgate.net/scientific-contributions/81760095_Stephen_R_Westrop |
| Siveter, D. J., Rushton, A. W. A., Siveter, D. J., 1995. An Ostracod-Like Arthropod with Appendages Preserved from the Lower Ordovician of England. Lethaia, 28(4): 299-307. doi: 10.1111/j.1502-3931.1995.tb01818.x |
| Siveter, D. J., Tanaka, G., Farrell, Ú. C., et al., 2014. Exceptionally Preserved 450-Million-Year-Old Ordovician Ostracods with Brood Care. Current Biology, 24(7): 801-806. doi: 10.1016/j.cub.2014.02.040 |
| Song, Y. Y., Zhang, Y. D., Zhang, J., 2013. New Advance in the Late Darriwillian to Early Sandbian Graptolite Biostratigraphy of Western Zhejiang and Northeastern Jiangxi Province, SE China. Journal of Stratigraphy, 37(2): 144-154 (in Chinese with English Abstract) |
| Spivey, R. C., 1939. Ostracodes from the Maquoketa Shale, Upper Ordovician, of Iowa. Journal of Paleontology, 13(2): 163-175 http://jpaleontol.geoscienceworld.org/content/13/2/163 |
| Sun, Q. Y., 1988. Ordovician Ostracoda from Western Hubei. Acta Micropalaeontologica Sinica, 5(3): 253-266 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-GSWX196104005.htm |
| Swain, F. M., 1957. Early Middle Ordovician Ostracoda of the Eastern United States. Part Ⅰ. Stratigraphic Data and Description of Leperditiidae, Aparchitidae and Leperditellidae. Journal of Paleontology, 31(3): 528-570 |
| Swain, F. M., 1962. Early Middle Ordovician Ostracoda of the Eastern United States. Part Ⅱ. Leperditellacea, Hollinacea, Kloedenellacea, Bairdiacea and Superfamily Uncertain. Journal of Paleontology, 36(4): 719-744 |
| Tinn, O., Meidla, T., 1999. Ordovician Ostracodes from the Komstad Limestone. Bulletin of the Geological Society of Denmark, 46(1): 25-30 https://www.researchgate.net/profile/Oive_Tinn/publication/238094997_Ordovician_ostracodes_from_the_Komstad_Limestone/links/004635318c19032181000000.pdf?origin=publication_detail |
| Tinn, O., Meidla, T., 2001. Middle Ordovician Ostracods from the Lanna and Holen Limestones, South-Central Sweden. GFF, 123(3): 129-136. doi: 10.1080/11035890101233129 |
| Tinn, O., Meidla, T., 2003. Ontogeny and Thanatocoenoses of Early Middle Ordovician Palaeocope Ostracode Species Brezelina Palmata (Krause, 1889) and Ogmoopsis Bocki (Öpik, 1935). Journal of Paleontology, 77(1): 64-72. doi: 10.1666/0022-3360(2003)077 |
| Umeda, M., Goto, H., Ishiga, H., 1992. Middle Ordovician Radiolarians from the Lachlan Fold Belt, Southeastern Australia. Memoirs of the Faculty of Science, Shimane University, 26: 131-140 https://www.researchgate.net/publication/229995899_Late_Ordovician_stratigraphy_zircon_provenance_and_tectonics_Lachlan_Fold_Belt_southeastern_Australia |
| Vannier, J. M. C., Racheboeuf, P. R., Benedetto, J. L., 1995. Silurian-Early Devonian Ostracodes from South America (Argentina, Bolivia): Preliminary Investigations.Journal of Paleontology, 69(4): 752-772. doi: 10.1017/s0022336000035265 |
| Wang, S. Q., 1988. Late Paleozoix Ostracode Association from South China and Their Paleoecological Significance. Acta Palaeontologica Sinica, 27: 91-102 (in Chinese with English Abstract) |
| Wang, S. Q., 2015. Ordovician and Silurian Ostracoda of China. China University of Technology Press, Hefei. 1-272 (in Chinese) |
| Wang, X. F., 1989. Palaeogeographic Reconstruction of Ordovician in China and Characteristics of Its Sedimentary Environment and Biofacies. Acta Micropalaeontologica Sinica, 28(2): 234-248 (in Chinese with English Abstract) http://edu.wanfangdata.com.cn/Periodical/Detail/gdlxb200802007 |
| Wang, X. F., 2016. Ordovician Tectonic-Paleogeography in South China and Chrono- and Bio-Stratigraphic Division and Correlation. Earth Science Prontiers, 23(6): 253-267 (in Chinese with English Abstract) https://www.researchgate.net/publication/251063198_Chrono_meteo_bio |
| Wang, Y. J., 1991. On Progress in the Study of Paleozoic Radiolarians in China. Acta Micropalaeontologica Sinica, 8(3): 237-251 (in Chinese with English Abstract) https://www.researchgate.net/profile/Jonathan_Aitchison/publication/238946738_Devonian_radiolarian_biostratigraphy_of_Guangxi_Province_SW_China/links/00b7d51c11e2cf116f000000.pdf?inViewer=true&pdfJsDownload=true&disableCoverPage=true&origin=publication_detail |
| Wang, Y. J., 1993. Middle Ordovician Radiolarians from the Pingliang Formation of Gansu Province, China. Micropaleontology Special Publication, 6: 98-114 http://www.irgrid.ac.cn/handle/1471x/752069 |
| Wang, Y. J., Cheng, J. F., Zhang, Y. D., 2008. New Radiolarian Genera and Species of Heituao Formation (Ordovician) in the Kuruktag Region, Xinjiang. Acta Palaeontologica Sinica, 47(4): 393-404 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTotal-GSWX200804003.htm |
| Wang, Y. J., Zhang, Y. D., 2011. Radiolarian Fauna of the Wufeng Formation (Upper Ordovician) in Lunshan Area, Jiangsu and Its Geological Significance. Acta Micropalaeontologica Sinica, 28(3): 251-260 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-WSGT201103001.htm |
| Wang, Z. H., Bergstrom, S. M., Zhang, Y. D., 2015. Upper Ordovician Conodonts from the Yenwashan Formation in the Zhejiang-Jiangxi Border Region, S.E. China and Their Biostratigraphic Significance. Acta Palaeontologica Sinica, 54: 147-157 |
| Williams, M., Stone, P., Siveter, D. J., et al., 2001. Upper Ordovician Ostracods from the Cautley District, Northern England: Baltic and Laurentian Affinities. Geological Magazine, 138(5): 589-607. doi: 10.1017/s0016756801005726 |
| Williams, M., Siveter, D. J., 1996. Lithofacies-Influenced Ostracod Associations in the Middle Ordovician Bromide Formation, Oklahoma, USA. Journal of Micropalaeontology, 15(1): 69-81. doi: 10.1144/jm.15.1.69 |
| Won, M. Z., Iams, W. J., 2011. Earliest Arenig Radiolarians from the Cow Head Group, Western Newfoundland. Journal of Paleontology, 85(1): 156-177. doi: 10.1666/10-102.1 |
| Won, M. Z., Iams, W. J., 2013. Early Ordovician (Early Arenig) Radiolarians from the Cow Head Group and Review of the Little Port Complex Fauna, Western Newfoundland. Palaeoworld, 22(1/2): 10-31. doi: 10.1016/j.palwor.2012.11.001 |
| Won, M. Z., Iams, W. J., 2015a. Early/Middle Arenig (Late Floian) Radiolarian Faunal Assemblages from Cow Head Group, Western Newfoundland. Palaeontographica Abteilung A, 304(1/2/3/4/5/6): 1-63. doi: 10.1127/pala/304/2015/1 |
| Won, M. Z., Iams, W. J., 2015b. Review of the Beothuka Terranova Assemblage and Characteristics of the Middle Arenig (Ordovician, Latest Floian) Radiolarian Assemblage from the Cow Head Group, Newfoundland. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 278(1): 1-21. doi: 10.1127/njgpa/2015/0513 |
| Wu, R. C., Stouge, S., Wang, Z. H., 2012. Conodontophorid Biodiversification during the Ordovician in South China. Lethaia, 45(3): 432-442. doi: 10.1111/j.1502-3931.2011.00303.x |
| Yan, K., Servais, T., Li, J., et al., 2011. Biodiversity Patterns of Early- Middle Ordovician Marine Microphytoplankton in South China. Palaeogeography, Palaeoclimatology, Palaeoecology, 299(1/2): 318-334. doi: 10.1016/j.palaeo.2010.11.012 |
| Yuan, F. T., Ma, L. X., 1993. Middle Ordovician Ostracodes from the Baota Formation Liangshan, Shanxi. Acta Micropalaeontologica Sinica, 10(2): 223-235 (in Chinese with English Abstract) |
| Zhan, R. B., Harper, D., 2006. Biotic Diachroneity during the Ordovician Radiation: Evidence from South China. Lethaia, 39(3): 211-226. doi: 10.1080/00241160600799770 |
| Zhan, R. B., Jin, J., 2007. Ordovician-Early Silurian (Llandovery) Stratigraphy and Palaeontology of the Upper Yangtze Platform, South China. Science Press, Beijing. 1-165 (in Chinese with English Abstract) |
| Zhan, R. B., Jin, J. S., Liu, J. B., 2013. Investigation on the Great Ordovician Biodiversification Event (GOBE): Review and Prospect. Chinese Science Bulletin (Chinese Version), 58(33): 3357-3371. doi:10.1360/972013-19 (in Chinese) |
| Zhan, R. B., Rong, J. Y., Cheng, J. H., et al., 2005. Early-Mid Ordovician Brachiopod Diversification in South China. Science in China Series D: Earth Sciences, 48(5): 662-675. doi: 10.1360/03yd0586 |
| Zhan, R. B., Zhang, Y. D., Yuan, W. W., 2007. A New Concept during the Life Process of the Earth-The Great Ordovician Biodiversification Event. Natural Science Process, 17(8): 1006-1014 (in Chinese with English Abstract) |
| Zhang, Y. D., Chen, X., 2008. The Evolution of Diversity and Environmental Background of the Ordovician Graptolites. Science in China Series D: Earth Sciences, 38(1): 10-21 (in Chinese) https://www.researchgate.net/profile/Yuandong_Zhang/publication/226046064_Diversity_history_of_Ordovician_graptolites_and_its_relationship_with_environmental_change/links/00b495295e5a60bd20000000.pdf?inViewer=true&disableCoverPage=true&origin=publication_detail |
| Zhang, Y. D., Chen, X., Goldman, D., 2007. Diversification Patterns of Early and Mid Ordovician Graptolites in South China. Geological Journal, 42(3/4): 315-337. doi: 10.1002/gj.1075 |
| Zhang, Y. D., Zhan, R. B., Fan, J. X., et al., 2009. The Key Scientific Problems of the Research on the Ordovician Radiation. Science in China Series D: Earth Sciences, 39(2): 129-143 (in Chinese) https://www.researchgate.net |
| Zhen, Y. Y., Zhou, Z. Y., 2008. Trilobite Record of China. Science Press, Beijing. 301-330 (in Chinese) |
| Zheng, N., Song, T. R., Li, T. D., et al., 2012. The Discovery of the Lower Cambrian and Middle Ordovician Radiolaria in the South China Orogenic Belt. Geology in China, 39(1): 260-265 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTotal-DIZI201201027.htm |
| Zhou, Z. Y., Yuan, J. L., Zhang, Z. H., et al., 1979. Cambrian Biogeographical Province of Guizhou and the Neighbouring Areas. Journal of Stratigraphy, 4(4): 258-271 (in Chinese) doi: 10.1007/978-3-642-79520-6_5 |
| Zhou, Z. Y., Yuan, W. W., Zhou, Z. Q., 2007. Patterns, Processes and Likely Causes of the Ordovician Trilobite Radiation in South China. Geological Journal, 42(3/4): 297-313. doi: 10.1002/gj.1076 |
| Zhou, Z. Y., Zhen, Y. Y., Zhou, Z. Q., 2008. Division of the Ordovician Geographic Untis of China—A Synopsis. Journal of Palaeogeography, 10(2): 176-182 (in Chinese with English Abstract) https://www.britannica.com/science/Ordovician-Period |
| Zong, R. W., Wang, Z. Z., Gong, Y. M., et al., 2015. Ordovician Radiolarians from the Yinisala Ophiolitic Mélange and Their Significance in Western Junggar, Xinjiang, NW China. Science China Earth Sciences, 58(5): 776-783. doi: 10.1007/s11430-014-4974-5 |
Figures(5) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: