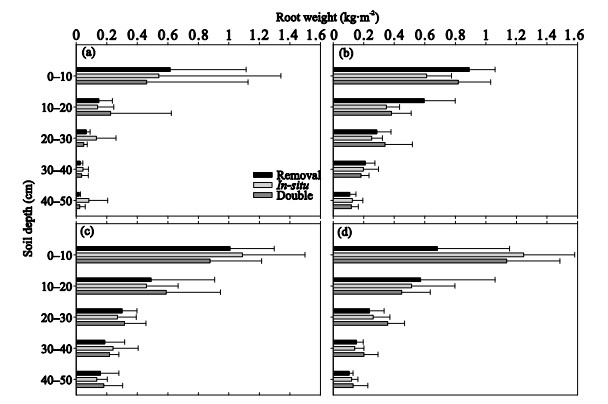
| Citation: | Yunwu Xiong, Bing Yu, Mengting Bai, Xueyang Zhang, Guanhua Huang, Alex Furman. Soil Properties and Plant Growth Response to Litter in a Prolonged Enclosed Grassland of Loess Plateau, China. Journal of Earth Science, 2019, 30(5): 1041-1048. doi: 10.1007/s12583-019-1017-3

|

| Baer, S. G., Blair, J. M., 2008. Grassland Establishment under Varying Resource Availability: A Test of Positive and Negative Feedback. Ecology, 89(7): 1859–1871. https://doi.org/10.1890/07-0417.1 |
| Ball, B. A., Carrillo, Y., Molina, M., 2014. The Influence of Litter Composition across the Litter-Soil Interface on Mass Loss, Nitrogen Dynamics and the Decomposer Community. Soil Biology and Biochemistry, 69: 71–82. https://doi.org/10.1016/j.soilbio.2013.10.048 |
| Bansal, S., Sheley, R. L., Blank, B., et al., 2014. Plant Litter Effects on Soil Nutrient Availability and Vegetation Dynamics: Changes that Occur When Annual Grasses Invade Shrub-Steppe Communities. Plant Ecology, 215(3): 367–378. https://doi.org/10.1007/s11258-014-0307-1 |
| Boeken, B., Orenstein, D., 2001. The Effect of Plant Litter on Ecosystem Properties in a Mediterranean Semi-Arid Shrubland. Journal of Vegetation Science, 12(6): 825–832. https://doi.org/10.2307/3236870 |
| Borgen, S. K., Molstad, L., Bruun, S., et al., 2011. Estimation of Plant Litter Pools and Decomposition-Related Parameters in a Mechanistic Model. Plant and Soil, 338(1/2): 205–222. https://doi.org/10.1007/s11104-010-0404-4 |
| Carrera, A. L., Bertiller, M. B., 2010. Relationships among Plant Litter, Fine Roots, and Soil Organic C and N Across an Aridity Gradient in Northern Patagonia, Argentina. coscience, 17(3): 276–286. https://doi.org/10.2980/17-3-3359 |
| Carrillo, Y., Ball, B. A., Strickland, M. S., et al., 2012. Legacies of Plant Litter on Carbon and Nitrogen Dynamics and the Role of the Soil Community. Pedobiologia, 55(4): 185–192. https://doi.org/10.1016/j.pedobi.2012.02.002 |
| Chang, X. F., Chai, Q. L., Wu, G. L., et al., 2017. Soil Organic Carbon Accumulation in Abandoned Croplands on the Loess Plateau. Land Degradation & Development, 28(5): 1519–1527. https://doi.org/10.1002/ldr.2679 |
| Chen, Y. P., Wang, K. B., Lin, Y. S., et al., 2015. Balancing Green and Grain Trade. Nature Geoscience, 8(10): 739–741. https://doi.org/10.1038/ngeo2544 |
| Deutsch, E. S., Bork, E. W., Willms, W. D., 2010. Soil Moisture and Plant Growth Responses to Litter and Defoliation Impacts in Parkland Grasslands. Agriculture, Ecosystems & Environment, 135(1/2): 1–9. https://doi.org/10.1016/j.agee.2009.08.002 |
| Dormaar, J. F., Adams, B. W., Willms, W. D., 1997. Impacts of Rotational Grazing on Mixed Prairie Soils and Vegetation. Journal of Range Management, 50(6): 647–651. https://doi.org/10.2307/4003461 |
| Facelli, J. M., Pickett, S. T. A., 1991a. Plant Litter: Its Dynamics and Effects on Plant Community Structure. The Botanical Review, 57(1): 1–32. https://doi.org/10.1007/bf02858763 |
| Facelli, J. M., Pickett, S. T. A., 1991b. Plant Litter: Light Interception and Effects on an Old-Field Plant Community. Ecology, 72(3): 1024–1031. https://doi.org/10.2307/1940602 |
| Freschet, G. T., Cornwell, W. K., Wardle, D. A., et al., 2013. Linking Litter Decomposition of Above- and Below-Ground Organs to Plant-Soil Feedbacks Worldwide. Journal of Ecology, 101(4): 943–952. https://doi.org/10.1111/1365-2745.12092 |
| Fu, B. J., Liu, Y., Lü, Y. H., et al., 2011. Assessing the Soil Erosion Control Service of Ecosystems Change in the Loess Plateau of China. Ecological Complexity, 8(4): 284–293. https://doi.org/10.1016/j.ecocom.2011.07.003 |
| Gao, Y., Cheng, J. M., 2013. Spatial and Temporal Variations of Grassland Soil Organic Carbon and Total Nitrogen Following Grazing Exclusion in Semiarid Loess Plateau, Northwest China. Acta Agriculturae Scandinavica, Section B: Soil & Plant Science, 63(8): 704–711. https://doi.org/10.1080/09064710.2013.854828 |
| Guretzky, J. A., Schacht, W. H., Wingeyer, A., et al., 2014. Litter Deposition and Nitrogen Return in Rotationally Stocked Smooth Bromegrass Pastures. Agronomy Journal, 106(1): 175–184. https://doi.org/10.2134/agronj2013.0282 |
| Horton, R., Kluitenberg, G., Vristow, K., et al., 1994. Surface Crop Residue Effects on the Soil Surface Energy Balance. Managing Agricultural Residues, 1: 143–162 http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=bb642a4a0ce3cbc45c50bc1bbfe44240 |
| Knapp, A. K., Seastedt, T. R., 1986. Detritus Accumulation Limits Productivity of Tallgrass Prairie. BioScience, 36(10): 662–668. https://doi.org/10.2307/1310387 |
| Leff, J. W., Wieder, W. R., Taylor, P. G., et al., 2012. Experimental Litterfall Manipulation Drives Large and Rapid Changes in Soil Carbon Cycling in a Wet Tropical Forest. Global Change Biology, 18(9): 2969–2979. https://doi.org/10.1111/j.1365-2486.2012.02749.x |
| Li, J. P., Zheng, Z. R., Xie, H. T., et al., 2017. Increased Soil Nutrition and Decreased Light Intensity Drive Species Loss after Eight Years Grassland Enclosures. Scientific Reports, 7(1): 44525. https://doi.org/10.1038/srep44525 |
| Liu, W. G., Wei, J., Cheng, J. M., et al., 2014. Profile Distribution of Soil Inorganic Carbon along a Chronosequence of Grassland Restoration on a 22-Year Scale in the Chinese Loess Plateau. Catena, 121: 321–329. https://doi.org/10.1016/j.catena.2014.05.019 |
| Naeth, M. A., Bailey, A. W., Chanasyk, D. S., et al., 1991. Water Holding Capacity of Litter and Soil Organic Matter in Mixed Prairie and Fescue Grassland Ecosystems of Alberta. Journal of Range Management, 44(1): 13–17. https://doi.org/10.2307/4002630 |
| Qiu, L. P., Wei, X. R., Zhang, X. C., et al., 2012. Soil Organic Carbon Losses Due to Land Use Change in a Semiarid Grassland. Plant and Soil, 355(1/2): 299–309. https://doi.org/10.1007/s11104-011-1099-x |
| Sakaguchi, K., Zeng, X. B., 2009. Effects of Soil Wetness, Plant Litter, and Under-Canopy Atmospheric Stability on Ground Evaporation in the Community Land Model (CLM3.5). Journal of Geophysical Research: Atmospheres, 114(D1): 010834. https://doi.org/10.1029/2008jd010834 |
| Sayer, E. J., 2006. Using Experimental Manipulation to Assess the Roles of Leaf Litter in the Functioning of Forest Ecosystems. Biological Reviews, 81(1): 1–31. https://doi.org/10.1017/s1464793105006846 |
| Villalobos-Vega, R., Goldstein, G., Haridasan, M., et al., 2011. Leaf Litter Manipulations Alter Soil Physicochemical Properties and Tree Growth in a Neotropical Savanna. Plant and Soil, 346(1/2): 385–397. https://doi.org/10.1007/s11104-011-0860-5 |
| Wang, J., Zhao, M. L., Willms, W. D., et al., 2011. Can Plant Litter Affect Net Primary Production of a Typical Steppe in Inner Mongolia?. Journal of Vegetation Science, 22(2): 367–376. https://doi.org/10.1111/j.1654-1103.2011.01257.x |
| Weber, K. T., Gokhale, B. S., 2011. Effect of Grazing on Soil-Water Content in Semiarid Rangelands of Southeast Idaho. Journal of Arid Environments, 75(5): 464–470. https://doi.org/10.1016/j.jaridenv.2010.12.009 |
| Wei, J., Liu, W. G., Cheng, J. M., et al., 2011. Dynamics of Soil Organic Carbon Storage Following Restoration of Grassland on Yunwu Mountain. Acta Ecologica Sinica, 31(5): 271–275. https://doi.org/10.1016/j.chnaes.2011.06.009 |
| Wickings, K., Grandy, A. S., Reed, S. C., et al., 2012. The Origin of Litter Chemical Complexity during Decomposition. Ecology Letters, 15(10): 1180–1188. https://doi.org/10.1111/j.1461-0248.2012.01837.x |
| Willms, W. D., McGinn, S. M., Dormaar, J. F., 1993. Influence of Litter on Herbage Production in the Mixed Prairie. Journal of Range Management, 46(4): 320–324. https://doi.org/10.2307/4002466 |
| Xiong, S. J., Nilsson, C., 1999. The Effects of Plant Litter on Vegetation: A Meta-Analysis. Journal of Ecology, 87(6): 984–994. https://doi.org/10.1046/j.1365-2745.1999.00414.x |
| Xu, S., Liu, L. L., Sayer, E. J., 2013. Variability of Above-Ground Litter Inputs Alters Soil Physicochemical and Biological Processes: A Meta-Analysis of Litterfall-Manipulation Experiments. Biogeosciences, 10(11): 7423–7433. https://doi.org/10.5194/bg-10-7423-2013 |
Figures(6) / Tables(3)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: