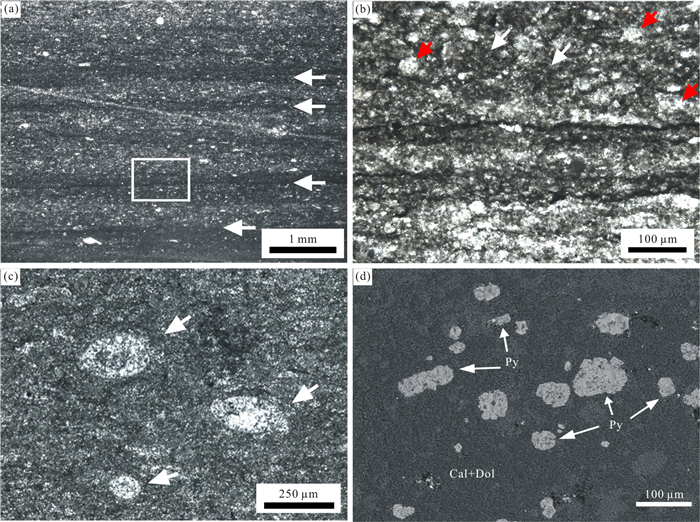
| Citation: | Mao Luo, Jitao Chen, Wenkun Qie, Jinyuan Huang, Qiyue Zhang, Changyong Zhou, Wen Wen. Microbially Induced Carbonate Precipitation in a Middle Triassic Microbial Mat Deposit from Southwestern China: New Implications for the Formational Process of Micrite. Journal of Earth Science, 2021, 32(3): 633-645. doi: 10.1007/s12583-020-1075-6

|

| Allwood, A. C., Walter, M. R., Kamber, B. S., et al., 2006. Stromatolite Reef from the Early Archaean Era of Australia. Nature, 441(7094): 714-718. https://doi.org/10.1038/nature04764 |
| Aloisi, G., Gloter, A., Krüger, M., et al., 2006. Nucleation of Calcium Carbonate on Bacterial Nanoglobules. Geology, 34(12): 1017-1020. https://doi.org/10.1130/g22986a.1 |
| Arp, G., Reimer, A., Reitner, J., 2003. Microbialite Formation in Seawater of Increased Alkalinity, Satonda Crater Lake, Indonesia. Journal of Sedimentary Research, 73(1): 105-127. https://doi.org/10.1306/071002730105 |
| Bai, J. K., Yin, F. G., Zhang, Q. Y., 2011. Microfacies and Enrichment Pattern of Fossils in the Fossiliferous Beds of Luoping Biota, Yunnan Province. Geology in China, 38: 393-402 (in Chinese with English Abstract) |
| Baumgartner, L. K., Reid, R. P., Dupraz, C., et al., 2006. Sulfate Reducing Bacteria in Microbial Mats: Changing Paradigms, New Discoveries. Sedimentary Geology, 185(3/4): 131-145. https://doi.org/10.1016/j.sedgeo.2005.12.008 |
| Benton, M. J., Zhang, Q. Y., Hu, S. X., et al., 2013. Exceptional Vertebrate Biotas from the Triassic of China, and the Expansion of Marine Ecosystems after the Permo-Triassic Mass Extinction. Earth-Science Reviews, 125: 199-243. https://doi.org/10.1016/j.earscirev.2013.05.014 |
| Berkyová, S., Munnecke, A., 2010. "Calcispheres" as a Source of Lime Mud and Peloids Evidence from the Early Middle Devonian of the Prague Basin, the Czech Republic. Bulletin of Geosciences, 85(4): 585-602. https://doi.org/10.3140/bull.geosci.1206 |
| Blue, C. R., Giuffre, A., Mergelsberg, S., et al., 2017. Chemical and Physical Controls on the Transformation of Amorphous Calcium Carbonate into Crystalline CaCO3 Polymorphs. Geochimica et Cosmochimica Acta, 196: 179-196. https://doi.org/10.1016/j.gca.2016.09.004 |
| Bontognali, T. R. R., Vasconcelos, C., Warthmann, R. J., et al., 2008. Microbes Produce Nanobacteria-Like Structures, Avoiding Cell Entombment. Geology, 36(8): 663-666. https://doi.org/10.1130/g24755a.1 |
| Bosak, T., Newman, D. K., 2003. Microbial Nucleation of Calcium Carbonate in the Precambrian. Geology, 31(7): 577-580. https://doi.org/10.1130/0091-7613(2003)031<0577:mnocci>2.0.co;2 doi: 10.1130/0091-7613(2003)031<0577:mnocci>2.0.co;2 |
| Braissant, O., Cailleau, G., Dupraz, C., et al., 2003. Bacterially Induced Mineralization of Calcium Carbonate in Terrestrial Environments: The Role of Exopolysaccharides and Amino-Acids. Journal of Sedimentary Research, 73(3): 483-488. https://doi.org/10.1306/111302730485 |
| Braissant, O., Decho, A. W., Dupraz, C., et al., 2007. Exopolymeric Substances of Sulfate-Reducing Bacteria: Interactions with Calcium at Alkaline pH and Implication for Formation of Carbonate Minerals. Geobiology, 5(4): 401-411. https://doi.org/10.1111/j.1472-4669.2007.00117.x |
| Bradley, J. P., Harvey, R. P., McSween, H. Y. Jr., 1997. No 'Nanofossils' in Martian Meteorite. Nature, 390: 454-456. https://doi.org/10.1038/37257 |
| Branda, S. S., Vik, A., Friedman, L., et al., 2005. Biofilm: The Matrix Revisited. Trends in Microbiology, 13(1): 20-26. https://doi.org/10.1016/j.tim.2004.11.006 |
| Broecker, S. W., Sanyal, A., Takahashi, T., 2000. The Origin of Bahamian Whitings Revisited. Geophysical Research Letters, 27: 3759-3760. https://doi.org/10.1029/2000gl011872 |
| Buczynski, C., Chafetz, H. S., 1991. Habit of Bacterially Induced Precipitates of Calcium Carbonate and the Influence of Medium Viscosity on Mineralogy. Journal of Sedimentary Petrology, 61(2): 226-233 doi: 10.1306/D42676DB-2B26-11D7-8648000102C1865D |
| Cai, Y. P., Xiao, S. H., Li, G. X., et al., 2019. Diverse Biomineralizing Animals in the Terminal Ediacaran Period Herald the Cambrian Explosion. Geology, 47: 380-384. https://doi.org/10.1130/g45949.1 |
| Cantine, M. D., Knoll, A. H., Bergmann, K. D., 2019. Carbonate before Skeletons: A Database Approach. Earth-Science Reviews, 201: 103065. https://doi.org/10.1016/j.earscirev.2019.103065 |
| Chafetz, H. S., Buczynski, C., 1992. Bacterially Induced Lithification of Microbialmats. Palaios, 7: 277-293. https://doi.org/10.2307/3514973 |
| Chen, Z. -Q., Benton, M. J., 2012. The Timing and Pattern of Biotic Recovery Following the End-Permian Mass Extinction. Nature Geoscience, 5: 375-383. https://doi.org/10.1038/ngeo1475 |
| Chen, Z. -Q., Wang, Y. B., Kershaw, S., et al., 2014. Early Triassic Stromatolites in a Siliciclastic Nearshore Setting in Northern Perth Basin, Western Australia: Geobiologic Features and Implications for Post-Extinction Microbial Proliferation. Global and Planetary Change, 121: 89-100. https://doi.org/10.1016/j.gloplacha.2014.07.004 |
| Cuadrado, D. G., Pan, J., 2018. Field Observations on the Evolution of Reticulate Patterns in Microbial Mats in a Modern Siliciclastic Coastal Environment. Journal of Sedimentary Research, 88(1): 24-37. https://doi.org/10.2110/jsr.2017.79 |
| Debenay, J. P., Andre, J. P., Lesourd, M., 1999. Production of Lime Mud by Breakdown of Foraminiferal Tests. Marine Geology, 157: 159-170. https://doi.org/10.1016/s0025-3227(98)00151-0 |
| Dimasi, E., Kwak, S. Y., Amos, F. F., 2006. Complementary Control by Additives of the Kinetics of Amorphous CaCO3 Mineralization at an Organic Interface: In-situ Synchrotron X-Ray Observations. Physical Review Letters, 97(4): 045503. https://doi.org/10.1103/physrevlett.97.045503 |
| Dupraz, C., Reid, R. P., Braissant, O., et al., 2009. Process of Carbonate Precipitation in Modern Microbial Mats. Earth-Science Reviews, 96(3): 141-162. https://doi.org/10.1016/j.earscirev.2008.10.005 |
| Dupraz, C., Visscher, P. T., 2005. Microbial Lithification in Marine Stromatolites and Hypersaline Mats. Trends in Microbiology, 13(9): 429-438. https://doi.org/10.1016/j.tim.2005.07.008 |
| Dupraz, C., Visscher, P. T., Baumgartner, L. K., et al., 2004. Microbe-Mineral Interaction: Early Carbonate Precipitation in a Hypersaline Lake (Eleuthera Island, Bahamas). Sedimentology, 51(4): 745-765. https://doi.org/10.1111/j.1365-3091.2004.00649.x |
| Enos, P., Lehrmann, D. J., Wei, J. Y., et al., 2006. Triassic Evolution of the Yangtze Platform in Guizhou Province, People's Republic of China. Geological Society of America Special Papers, 417: 1-105. https://doi.org/10.1130/spe417 |
| Enríquez, S., Schubert, N., 2014. Direct Contribution of the Seagrass Thalassiatestudinum to Lime Mud Production. Nature Communications, 5: e3835. https://doi.org/10.1038/ncomms4835 |
| Ercole, C., Cacchio, P., Botta, A. L., et al., 2007. Bacterially Induced Mineralization of Calcium Carbonate: The Role of Exopolysaccharides and Capsular Polysaccharides. Microscopy and Microanalysis, 13(1): 42-50. https://doi.org/10.1017/s1431927607070122 |
| Fang, Y. H., Chen, Z. Q., Kershaw, S., et al., 2017. Permian-Triassic Boundary Microbialites at Zuodeng Section, Guangxi Province, South China: Geobiology and Palaeoceanographic Implications. Global and Planetary Change, 152: 115-128. https://doi.org/10.1016/j.gloplacha.2017.02.011 |
| Feldmann, R. M., Schweitzer, C. E., Hu, S. X., et al., 2012. Macrurous Decapoda from the Luoping Biota (Middle Triassic) of China. Journal of Paleontology, 86(3): 425-441. https://doi.org/10.1666/11-113.1 |
| Feldmann, R. M., Schweitzer, C. E., Hu, S. X., et al., 2015. Spatial Distribution of Crustacea and Associated Organisms in the Luoping Biota (Anisian, Middle Triassic), Yunnan Province, China: Evidence of Periodic Mass Kills. Journal of Paleontology, 89(6): 1022-1037. https://doi.org/10.1017/jpa.2015.60 |
| Feldmann, R. M., Schweitzer, C. E., Hu, S. X., et al., 2017. A New Middle Triassic (Anisian) Cyclidan Crustacean from the Luoping Biota, Yunnan Province, China: Morphologic and Phylogenetic Insights. Journal of Crustacean Biology, 37(4): 406-412. https://doi.org/10.1093/jcbiol/rux052 |
| Feng, X. Q., Chen, Z. Q., Bottjer, D. J., et al., 2018. Additional Records of Ichnogenus Rhizocorallium from the Lower and Middle Triassic, South China: Implications for Biotic Recovery after the End-Permian Mass Extinction. GSA Bulletin, 130(7/8): 1197-1215. https://doi.org/10.1130/b31715.1 |
| Feng, X. Q., Chen, Z. Q., Woods, A., et al., 2017. Anisian (Middle Triassic) Marine Ichnocoenoses from the Eastern and Western Margins of the Kamdian Continent, Yunnan Province, SW China: Implications for the Triassic Biotic Recovery. Global and Planetary Change, 157: 194-213. https://doi.org/10.1016/j.gloplacha.2017.09.004 |
| Folk, R. L., Robles, R., 1964. Carbonate Sands of Isla Perez, Alacran Reef Complex, Yucatán. The Journal of Geology, 72(3): 255-292. https://doi.org/10.1086/626986 |
| Folk, R. L., 1993. SEM Imaging of Bacteria and Nannobacteria in Carbonate Sediments and Rocks. Journal of Sedimentary Petrology, 63: 990-999 |
| González-Muñoz, M. T., Rodriguez-Navarro, C., Martínez-Ruiz, F., et al., 2010. Bacterial Biomineralization: New Insights from Myxococcus-Induced Mineral Precipitation. Geological Society, London, Special Publications, 336(1): 31-50. https://doi.org/10.1144/sp336.3 |
| Grotzinger, J. P., Knoll, A. H., 1999. Stromatolites in Precambrian Carbonates: Evolutionary Mileposts or Environmental Dipsticks? Annual Review of Earth and Planetary Sciences, 27(1): 313-358. https://doi.org/10.1146/annurev.earth.27.1.313 |
| Hagadorn, J. W., Bottjer, D. J., 1997. Wrinkle Structures: Microbially Mediated Sedimentary Structures Common in Subtidal Siliciclastic Settings at the Proterozoic-Phanerozoic Transition. Geology, 25(11): 1047-1050. https://doi.org/10.1130/0091-7613(1997)025<1047:wsmmss>2.3.co;2 doi: 10.1130/0091-7613(1997)025<1047:wsmmss>2.3.co;2 |
| Han, Z. Z., Zhao, Y. Y., Yan, H. X., et al., 2017. The Characterization of Intracellular and Extracellular Biomineralization Induced by Synechocystis sp. PCC6803 Cultured under Low Mg/Ca Ratios Conditions. Geomicrobiology Journal, 34: 362-373. https://doi.org/10.1080/01490451.2016.1197986 |
| Hu, S. L., Li, Y. J., Dai, M., et al., 1996. The Laser Mass-Spectrometer 40Ar-49Ar Age of Green Pisolites of Guizhou Province. Acta Petrologica Sinica, 12: 409-415 (in Chinese with English Abstract) |
| Hu, S. X., Zhang, Q. Y., Chen, Z. -Q., et al., 2011. The Luoping Biota: Exceptional Preservation, and New Evidence on the Triassic Recovery from End-Permian Mass Extinction. Proceedings of the Royal Society B: Biological Sciences, 278(1716): 2274-2282. https://doi.org/10.1098/rspb.2010.2235 |
| Hu, S. X., Zhang, Q. Y., Feldmann, R. M., et al., 2017. Exceptional Appendage and Soft-Tissue Preservation in a Middle Triassic Horseshoe Crab from SW China. Scientific Reports, 7(1): 14112. https://doi.org/10.1038/s41598-017-13319-x |
| Hu, S. X., Zhang, Q. Y., Zhou, C. Y., 2010. Fossil Coprolites from the Middle Triassic Luoping Biota and Ecological Implication. Journal of Earth Science, 21(1): 191-193. https://doi.org/10.1007/s12583-010-0209-7 |
| Huang, J. Y., Hu, S. X., Zhang, Q. Y., et al., 2019a. Gondolelloid Multielement Conodont Apparatus (Nicoraella) from the Middle Triassic of Yunnan Province, Southwestern China. Palaeogeography, Palaeoclimatology, Palaeoecology, 522: 98-110. https://doi.org/10.1016/j.palaeo.2018.07.015 |
| Huang, J. Y., Martínez-Pérez, C., Hu, S. X., et al., 2019b. Apparatus Architecture of the Conodont Nicoraella kockeli (Gondolelloidea, Prioniodinina) Constrains Functional Interpretations. Palaeontology, 62(5): 823-835. https://doi.org/10.1111/pala.12429 |
| Huang, J. Y., Martínez-Pérez, C., Hu, S. X., et al., 2019c. Middle Triassic Conodont Apparatus Architecture Revealed by Synchrotron X-Ray Microtomography. Palaeoworld, 28(4): 429-440. https://doi.org/10.1016/j.palwor.2018.08.003 |
| Huang, J. Y., Zhang, K. X., Zhang, Q. Y., et al., 2009. Conodonts Stratigraphy and Sedimentary Environment of the Middle Triassic at Daaozi Section of Luoping County, Yunnan Province, South China. Acta Micropalaeontologica Sinica, 26(3): 211-224 (in Chinese with English Abstract) |
| Kawaguchi, T., Decho, A. W., 2002. Isolation and Biochemical Characterization of Extracellular Polymeric Secretions (EPS) from Modern Soft Marine Stromatolites (Bahamas) and Its Inhibitory Effect on CaCO3 Precipitation. Preparative Biochemistry & Biotechnology, 32(1): 51-63. https://doi.org/10.1081/pb-120013161 |
| Kaźmierczak, J., Coleman, M. L., Gruszczyński, M., et al., 1996. Cyanobacterial Key to the Genesis of Micritic and Peloidal Limestones in Ancient Seas. Acta Palaeontologica Polonica, 41(4): 319-338 |
| Kaźmierczak, J., Fenchel, T., Kühl, M., et al., 2015. CaCO3 Precipitation in Multilayered Cyanobacterial Mats: Clues to Explain the Alternation of Micrite and Sparite Layers in Calcareous Stromatolites. Life (Basel), 5(1): 744-769. https://doi.org/10.3390/life5010744 |
| Kirkland, B. L., Lynch, F. L., Rahnis, M. A., et al., 1999. Alternative Origins for Nannobacteria-Like Objects in Calcite. Geology, 27(4): 347-350. https://doi.org/10.1130/0091-7613(1999)0270347:aofnlo>2.3.co;2 doi: 10.1130/0091-7613(1999)0270347:aofnlo>2.3.co;2 |
| Knoll, A. H., Fairchild, I. J., Swett, K., 1993. Calcified Microbes in Neoproterozoic Carbonates: Implications for Our Understanding of the Proterozoic/Cambrian Transition. Palaios, 8: 512-525 doi: 10.2307/3515029 |
| Kremer, B., Kazmierczak, J., Stal, L. J., 2008. Calcium Carbonate Precipitation in Cyanobacterial Mats from Sandy Tidal Flats of the North Sea. Geobiology, 6(1): 46-56. https://doi.org/10.1111/j.1472-4669.2007.00128.x |
| Li, H., Yao, Q. Z., Wang, F. P., et al., 2019. Insights into the Formation Mechanism of Vaterite Mediated by a Deep-Sea Bacterium Shewanella piezotolerans WP3. Geochimica et Cosmochimica Acta, 256: 35-48. https://doi.org/10.1016/j.gca.2018.06.011 |
| Littlewood, J. L., Shaw, S., Peacock, C. L., et al., 2017. Mechanism of Enhanced Strontium Uptake into Calcite via an Amorphous Calcium Carbonate Crystallization Pathway. Crystal Growth & Design, 17(3): 1214-1223. https://doi.org/10.1021/acs.cgd.6b01599 |
| Lowenstam, H. A., 1955. Aragonite Needles Secreted by Algae and Some Sedimentary Implications. SEPM Journal of Sedimentary Research, 25: 270-272 |
| Luo, M., Chen, Z. -Q., Hu, S. X., et al., 2013. Carbonate Reticulated Ridge Structures from the Lower Middle Triassic of the Luoping Area, Yunnan, Southwestern China: Geobiologic Features and Implications for Exceptional Preservation of the Luoping Biota. PALAIOS, 28(8): 541-551. https://doi.org/10.2110/palo.2012.p12-122r |
| Luo, M., Chen, Z. -Q., Shi, G. -R., et al., 2016a. Upper Lower Triassic Stromatolite from Anhui, South China: Geobiologic Features and Paleoenvironmental Implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 452: 40-54. https://doi.org/10.1016/j.palaeo.2016.04.008 |
| Luo, M., Chen, Z. -Q., Zhao, L. S., et al., 2014. Early Middle Triassic Stromatolites from the Luoping Area, Yunnan Province, Southwest China: Geobiologic Features and Environmental Implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 412: 124-140. https://doi.org/10.1016/j.palaeo.2014.07.028 |
| Luo, M., George, A. D., Chen, Z. -Q., 2016b. Sedimentology and Ichnology of Two Lower Triassic Sections in South China: Implications for the Biotic Recovery Following the End-Permian Mass Extinction. Global and Planetary Change, 144: 198-212. https://doi.org/10.1016/j.gloplacha.2016.07.007 |
| Luo, M., Gong, Y. M., Shi, G. -R., et al., 2018. Palaeoecological Analysis of Trace Fossil Sinusichnus Sinuosus from the Middle Triassic Guanling Formation in Southwestern China. Journal of Earth Science, 29(4): 854-863. https://doi.org/10.1007/s12583-018-0794-4 |
| Luo, M., Hu, S. X., Benton, M. J., et al., 2017. Taphonomy and Palaeobiology of Early Middle Triassic Coprolites from the Luoping Biota, Southwest China: Implications for Reconstruction of Fossil Food Webs. Palaeogeography, Palaeoclimatology, Palaeoecology, 474: 232-246. https://doi.org/10.1016/j.palaeo.2016.06.001 |
| Luo, M., Shi, G. -R., Hu, S. X., et al., 2019. Early Middle Triassic Trace Fossils from the Luoping Biota, Southwestern China: Evidence of Recovery from Mass Extinction. Palaeogeography, Palaeoclimatology, Palaeoecology, 515: 6-22. https://doi.org/10.1016/j.palaeo.2017.11.028 |
| Nealson, K. H., Stahl, D. A., 1997. Microorganisms and Biogeochemical Cycles: What can we Learn from Layered Microbial Communities?. In: Banfield, J. F., Nealson, K. H., eds., Geomicrobiology, Interactions between Microbes and Minerals. Mineralogical Society of America, Washington, D.C. 35: 5-34. https: //doi.org/10.1515/9781501509247-003 |
| Nealson, K., 1999. Discussion. In: Steering, G., eds., Size Limit of Very Small Microorganisms: Proceedings of a Workshop. National Academy Press, National Research Council, Space studies Board, Washington, D.C. 39-42 |
| Pacton, M., Gorin, G., Vasconcelos, C., et al., 2010. Structural Arrangement of Sedimentary Organic Matter: Nanometer-Scale Spheroids as Evidence of a Microbial Signature in Early Diagenetic Processes. Journal of Sedimentary Research, 80(10): 919-932. https://doi.org/10.2110/jsr.2010.082 |
| Pratt, B. R., 2001. Calcification of Cyanobacterial Filaments: Girvanella and the Origin of Lower Paleozoic Lime Mud. Geology, 29(9): 763-766. https://doi.org/10.1130/0091-7613(2001)0290763:cocfga>2.0.co;2 doi: 10.1130/0091-7613(2001)0290763:cocfga>2.0.co;2 |
| Reid, R. P., Macintyre, I. G., 1992. Micritized Skeletal Grains in Northern Belize Lagoon: A Major Source of Mg-Calcite Mud. SEPM Journal of Sedimentary Research, 62: 145-156 |
| Riding, R., 2006. Cyanobacterial Calcification, Carbon Dioxide Concentrating Mechanisms, and Proterozoic-Cambrian Changes in Atmospheric Composition. Geobiology, 4(4): 299-316. https://doi.org/10.1111/j.1472-4669.2006.00087.x |
| Robbins, L. L., Blackwelder, P. L., 1992. Biochemical and Ultrastructural Evidence for the Origin of Whitings: A Biologically Induced Calcium Carbonate Precipitation Mechanism. Geology, 20(5): 464-468. https://doi.org/10.1130/0091-7613(1992)0200464:baueft>2.3.co;2 doi: 10.1130/0091-7613(1992)0200464:baueft>2.3.co;2 |
| Robbins, L. L., Tao, Y., Evans, C. A., 1997. Temporal and Spatial Distribution of Whitings on Great Bahama Bank and a New Lime Mud Budget. Geology, 25(10): 947-950. https://doi.org/10.1130/0091-7613(1997) 0250947:tasdow>2.3.co;2 doi: 10.1130/0091-7613(1997)0250947:tasdow>2.3.co;2 |
| Rodriguez-Blanco, J. D., Sand, K. K., Benning, L. G., 2017. ACC and Vaterite as Intermediates in the Solution-Based Crystallization of CaCO3. New Perspectives on Mineral Nucleation and Growth. Springer International Publishing, Cham. 93-111. https: //doi.org/10.1007/978-3-319-45669-0_5 |
| Rodriguez-Navarro, C., Jimenez-Lopez, C., Rodriguez-Navarro, A., et al., 2007. Bacterially Mediated Mineralization of Vaterite. Geochimica et Cosmochimica Acta, 71(5): 1197-1213. https://doi.org/10.1016/j.gca.2006.11.031 |
| Rule, R. G., Pratt, B. R., 2019. The Pseudofossil Horodyskia: Flocs and Flakes on Microbial Mats in a Shallow Mesoproterozoic Sea (Appekunny Formation, Belt Supergroup, Western North America). Precambrian Research, 333: 105439. https://doi.org/10.1016/j.precamres.2019.105439 |
| Sánchez-Román, M., Vasconcelos, C., Schmid, T., et al., 2008. Aerobic Microbial Dolomite at the Nanometer Scale: Implications for the Geological Record. Geology, 36(11): 879-882. https://doi.org/10.1130/g25013a.1 |
| Schieber, J., Arnott, H. J., 2003. Nannobacteria as a By-Product of Enzyme-Driven Tissue Decay. Geology, 31(8): 717-720. https://doi.org/10.1130/g19663.1 |
| Seong-Joo, L., Golubic, S., 1999. Microfossil Populations in the Context of Synsedimentary Micrite Deposition and Acicular Carbonate Precipitation: Mesoproterozoic Gaoyuzhuang Formation, China. Precambrian Research, 96(3/4): 183-208. https://doi.org/10.1016/s0301-9268(99)00004-2 |
| Spadafora, A., Perri, E., McKenzie, J. A., et al., 2010. Microbial Biomineralization Processes Forming Modern Ca: Mg Carbonate Stromatolites. Sedimentology, 57(1): 27-40. https://doi.org/10.1111/j.1365-3091.2009.01083.x |
| Stieglitz, R. D., 1972. Scanning Electron Microscopy of the Fine Fraction of Recent Carbonate Sediments from Bimini, Bahamas. Journal of Sedimentary Petrology, 37: 211-227 |
| Stockman, K. W., Ginsburg, R. N., Shinn, E. A., 1967. The Production of Lime Mud by Algae in South Florida. Journal of Sedimentary Petrology, 37: 633-648 |
| Sutherland, I. W., 2001. Microbial Polysaccharides from Gram-Negative Bacteria. International Dairy Journal, 11(9): 663-674. https://doi.org/10.1016/s0958-6946(01)00112-1 |
| Tang, D. J., Shi, X. Y., Jiang, G. Q., 2013. Mesoproterozoic Biogenic Thrombolites from the North China Platform. International Journal of Earth Sciences, 102(2): 401-413. https://doi.org/10.1007/s00531-012-0817-9 |
| Thompson, J. B., 2000. Microbial Whitings. In: Riding, R. E., Awramik, S. M., eds., Microbial Sediments. Springer, Berlin Heidelberg, New York. 250-260 |
| Tosti, F., Riding, R., 2017. Fine-Grained Agglutinated Elongate Columnar Stromatolites: Tieling Formation, ca. 1 420 Ma, North China. Sedimentology, 64(4): 871-902. https://doi.org/10.1111/sed.12336 |
| Trichet, J., Défarge, C., 1995. Non-Biologically Supported Organomineralization. I'Institut Océanographique de Monaco, Monaco. 203-236 |
| Trower, E. J., Lamb, M. P., Fischer, W. W., 2019. The Origin of Carbonate Mud. Geophysical Research Letters, 46(5): 2696-2703. https://doi.org/10.1029/2018gl081620 |
| Tucker, M. E., 2001. Sedimentary Petrology: An Introduction to the Origin of Sedimentary Rocks. Blackwell Science. 262 |
| Visscher, P. T., Reid, R. P., Bebout, B. M., et al., 1998. Formation of Lithified Micritic Laminae in Modern Marine Stromatolites (Bahamas); The Role of Sulfur Cycling. American Mineralogist, 83(11/12): 1482-1493. https://doi.org/10.2138/am-1998-11-1236 |
| Wen, W., Hu, S. X., Zhang, Q. Y., et al., 2019. A New Species of Platysiagum from the Luoping Biota (Anisian, Middle Triassic, Yunnan, South China) Reveals the Relationship between Platysiagidae and Neopterygii. Geological Magazine, 156(4): 669-682. https://doi.org/10.1017/s0016756818000079 |
| Wen, W., Zhang, Q. Y., Hu, S. X., et al., 2012. A New Basal Actinopterygian Fish from the Anisian (Middle Triassic) of Luoping, Yunnan Province, Southwest China. Acta Palaeontologica Polonica, 57(1): 149-160. https://doi.org/10.4202/app.2010.0089 |
| Wen, W., Zhang, Q. Y., Hu, S. X., et al., 2013. Coelacanths from the Middle Triassic Luoping Biota, Yunnan, South China, with the Earliest Evidence of Ovoviviparity. Acta Palaeontologica Polonica, 58(1): 175-193. https://doi.org/10.4202/app.2011.0066 |
| You, X. L., Sun, S., Zhu, J. Q., et al., 2013. Microbially Mediated Dolomite in Cambrian Stromatolites from the Tarim Basin, North-West China: Implications for the Role of Organic Substrate on Dolomite Precipitation. Terra Nova, 25(5): 387-395. https://doi.org/10.1111/ter.12048 |
| Zhang, Q. Y., Zhou, C. Y., Lu, T., et al., 2009. A Conodont-Based Middle Triassic Age Assignment for the Luoping Biota of Yunnan, China. Science in China Series D: Earth Sciences, 52(10): 1673-1678. https://doi.org/10.1007/s11430-009-0114-z |
Figures(10)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: