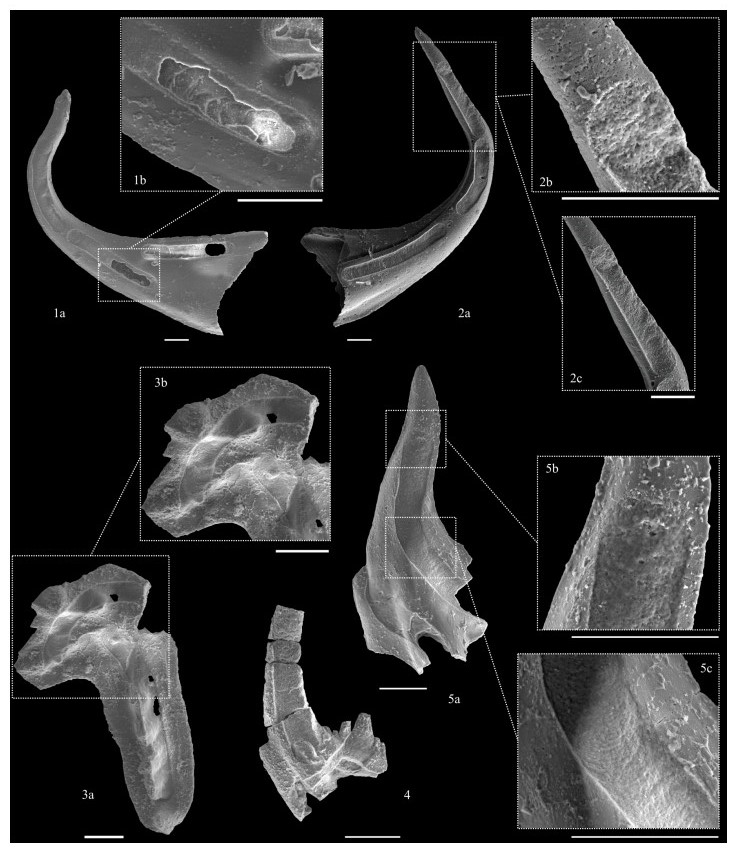
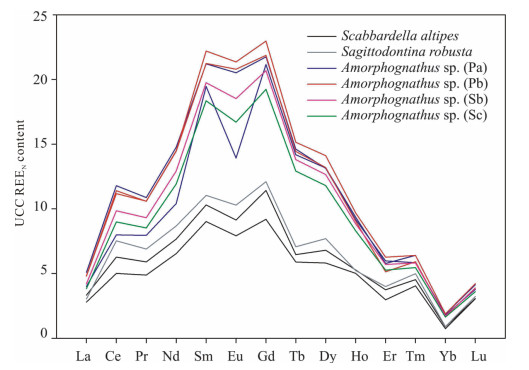
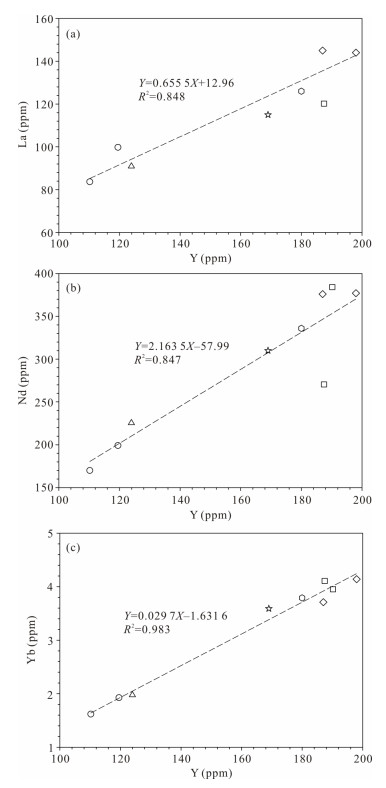
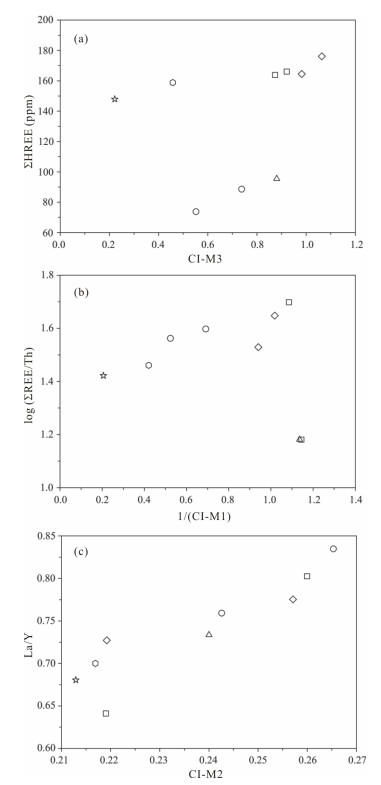
| Citation: | Luca Medici, Martina Savioli, Annalisa Ferretti, Daniele Malferrari. Zooming in REE and Other Trace Elements on Conodonts: Does Taxonomy Guide Diagenesis?. Journal of Earth Science, 2021, 32(3): 501-511. doi: 10.1007/s12583-020-1094-3

|

| Armstrong, H. A., Pearson, D. G., Griselin, M., 2001. Thermal Effects on Rare Earth Element and Strontium Isotope Chemistry in Single Conodont Elements. Geochimica et Cosmochimica Acta, 65(3): 435-441. https://doi.org/10.1016/s0016-7037(00)00548-2 doi: 10.1016/S0016-7037(00)00548-2 |
| Bergström, S. M., Ferretti, A., 2015. Conodonts in the Upper Ordovician Keisley Limestone of Northern England: Taxonomy, Biostratigraphical Significance and Biogeographical Relationships. Papers in Palaeontology, 1(1): 1-32. https://doi.org/10.1002/spp2.1003 |
| Bright, C. A., Cruse, A. M., Lyons, T. W., et al., 2009. Seawater Rare-Earth Element Patterns Preserved in Apatite of Pennsylvanian Conodonts?. Geochimica et Cosmochimica Acta, 73(6): 1609-1624. https://doi.org/10.1016/j.gca.2008.12.014 |
| Brigatti, M. F., Malferrari, D., Medici, L., et al., 2004. Crystal Chemistry of Apatites from the Tapira Carbonatite Complex, Brazil. European Journal of Mineralogy, 16(4): 677-685. https://doi.org/10.1127/0935-1221/2004/0016-0677 |
| Burnett, R. D., Hall, J. C., 1992. Significance of Ultrastructural Features in Etched Conodonts. Journal of Paleontology, 66(2): 266-276. https://doi.org/10.1017/s0022336000033783 doi: 10.1017/S0022336000033783 |
| Chen, J. B., Algeo, T. J., Zhao, L. S., et al., 2015. Diagenetic Uptake of Rare Earth Elements by Bioapatite, with an Example from Lower Triassic Conodonts of South China. Earth-Science Reviews, 149: 181-202. https://doi.org/10.1016/j.earscirev.2015.01.013 |
| Collins, M. J., Nielsen-Marsh, C. M., Hiller, J., et al., 2002. The Survival of Organic Matter in Bone: A Review. Archaeometry, 44(3): 383-394. https://doi.org/10.1111/1475-4754.t01-1-00071 |
| Cruse, A. M., Lyons, T. W., 2000. Sedimentology and Geochemistry of the Hushpuckney and Upper Tackett Shales Cyclothem Models Revisited. In: Johnson, K. S., ed., Marine Clastics in the Southern Midcontinent, 1997 Symposium. Oklahoma Geological Survey Circular, 103: 185-194 |
| Del Moral, B., Sarmiento, G. N., 2008. Conodontos del Katiense (Ordovicico Superior) del Sector Meridional de la Zone Centroibérica (España). Revista de Micropaleontologia, 40: 169-245 http://dialnet.unirioja.es/servlet/articulo?codigo=2945752 |
| Dzik, J., 1999. Evolution of Late Ordovician High-Latitude Conodonts and Dating of Gondwana Glaciations. Bollettino della Società Paleontologica Italiana, 37(2): 237-253 http://www.researchgate.net/publication/288394439_Evolution_of_the_Late_Ordovician_high-latitude_conodonts_and_dating_of_Gondwana_glaciations |
| Dzik, J., 2020. Ordovician Conodonts and the Tornquist Lineament. Palaeogeography, Palaeoclimatology, Palaeoecology, 549: 109157. https://doi.org/10.1016/j.palaeo.2019.04.013 |
| Ferretti, A., Bancroft, A. M., Repetski, J. E., 2020a. GECkO: Global Events Impacting Conodont Evolution. Palaeogeography, Palaeoclimatology, Palaeoecology, 549: 109677. https://doi.org/10.1016/j.palaeo.2020.109677 |
| Ferretti, A., Malferrari, D., Savioli, M., et al., 2020b. ʻConodont Pearlsʼ do not Belong to Conodonts. Lethaia. https://doi.org/10.1111/let.12403 |
| Ferretti, A., Barnes, C. R., 1997. Upper Ordovician Conodonts from the Kalkbank Limestone of Thuringia, Germany. Palaeontology, 40(1): 15-42 http://www.researchgate.net/publication/215580298_Upper_Ordovician_conodonts_from_the_Kalkbank_limestone_of_Thuringia_Germany |
| Ferretti, A., Bergström, S. M., Barnes, C. R., 2014a. Katian (Upper Ordovician) Conodonts from Wales. Palaeontology, 57(4): 801-831. https://doi.org/10.1111/pala.12089 |
| Ferretti, A., Bergström, S. M., Sevastopulo, G. D., 2014b. Katian Conodonts from the Portrane Limestone: The First Ordovician Conodont Fauna Described from Ireland. Bollettino della Società Paleontologica Italiana, 53(2): 105-119 http://www.researchgate.net/publication/266382330_Katian_conodonts_from_the_Portrane_Limestone_the_first_Ordovician_conodont_fauna_described_from_Ireland |
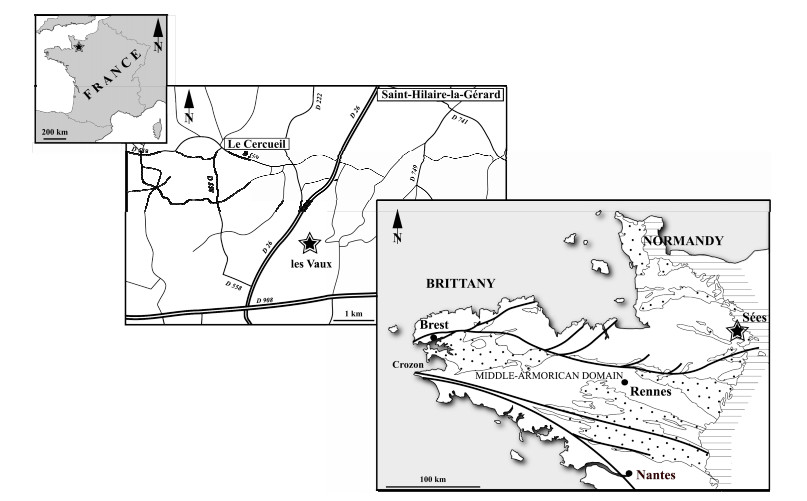
| Ferretti, A., Messori, A., Bergström, S. M., 2014c. Composition and Significance of the Katian (Upper Ordovician) Conodont Fauna of the Vaux Limestone ('Calcaire des Vaux') in Normandy, France. Estonian Journal of Earth Sciences, 63(4): 214-219. https://doi.org/10.3176/earth.2014.21 |
| Ferretti, A., Malferrari, D., Medici, L., et al., 2017. Diagenesis does not Invent anything New: Precise Replication of Conodont Structures by Secondary Apatite. Scientific Reports, 7(1): 1624. https://doi.org/10.1038/s41598-017-01694-4 |
| Ferretti, A., Schönlaub, H. P., 2001. New Conodont Faunas from the Late Ordovician of the Central Carnic Alps, Austria. Bollettino della Società Paleontologica Italiana, 40(1): 3-15 http://www.researchgate.net/publication/287005197_New_conodont_faunas_from_the_Late_Ordovician_of_the_Central_Carnic_Alps_Austria |
| Ferretti, A., Serpagli, E., 1991. First Record of Ordovician Conodonts from Southwestern Sardinia. Rivista Italiana di Paleontologia e Stratigrafia, 97(1): 27-34 http://www.researchgate.net/publication/291859932_First_record_of_Ordovician_conodonts_from_Southwestern_Sardinia |
| Ferretti, A., Serpagli, E., 1999. Late Ordovician Conodont Faunas from Southern Sardinia, Italy: Biostratigraphic and Paleogeographic Implications. Bollettino della Società Paleontologica Italiana, 37(2/3): 215-236 http://ci.nii.ac.jp/naid/10025919539 |
| Frank-Kamenetskaya, O. V., Rozhdestvenskaya, I. V., Rosseeva, E. V., et al., 2014. Refinement of Apatite Atomic Structure of Albid Tissue of Late Devon Conodont. Crystallography Reports, 59(1): 41-47. https://doi.org/10.1134/s1063774514010039 doi: 10.1134/S1063774514010039 |
| Girard, C., Albarède, F., 1996. Trace Elements in Conodont Phosphates from the Frasnian/Famennian Boundary. Palaeogeography, Palaeoclimatology, Palaeoecology, 126(1/2): 195-209. https://doi.org/10.1016/s0031-0182(96)00114-9 http://www.sciencedirect.com/science/article/pii/S0031018296001149 |
| Grandjean-Lécuyer, P., Feist, R., Albarède, F., 1993. Rare Earth Elements in Old Biogenic Apatites. Geochimica et Cosmochimica Acta, 57(11): 2507-2514. https://doi.org/10.1016/0016-7037(93)90413-q doi: 10.1016/0016-7037(93)90413-Q |
| Grandjean, P., Cappetta, H., Michard, A., et al., 1987. The Assessment of REE Patterns and 143Nd/144Nd Ratios in Fish Remains. Earth and Planetary Science Letters, 84(2/3): 181-196. https://doi.org/10.1016/0012-821x(87)90084-7 http://www.sciencedirect.com/science/article/pii/0012821X87900847 |
| Heckel, P. H., Baesemann, J. F., 1975. Environmental Interpretation of Conodont Distribution in Upper Pennsylvanian (Missourian) Megacyclothems in Eastern Kansas. AAPG Bulletin, 59: 486-509. https://doi.org/10.1306/83d91cb8-16c7-11d7-8645000102c1865d http://www.researchgate.net/publication/279601135_Environmental_interpretation_of_conodont_distribution_in_Upper_Pennsylvanian_Missourian_megacyclothems_in_Eastern_Kansas |
| Henningsmoen, G., 1948. The Tretaspis Series of the Kullatorp Core. In: Waern, B., Thorslund, P., Henningsmoen, G., eds., Deep Boring through Ordovician and Silurian Strata at Kinnekulle, Vestergötland. Bulletin of the Geological Institution of the University of Uppsala, 32: 374-432 |
| Herwartz, D., Tütken, T., Jochum, K. P., et al., 2013. Rare Earth Element Systematics of Fossil Bone Revealed by LA-ICPMS Analysis. Geochimica et Cosmochimica Acta, 103: 161-183. https://doi.org/10.1016/j.gca.2012.10.038 |
| Herwartz, D., Tütken, T., Münker, C., et al., 2011. Timescales and Mechanisms of REE and Hf Uptake in Fossil Bones. Geochimica et Cosmochimica Acta, 75(1): 82-105. https://doi.org/10.1016/j.gca.2010.09.036 |
| Holmden, C., Creaser, R. A., Muehlenbachs, K., et al., 1996. Isotopic and Elemental Systematics of Sr and Nd in 454 Ma Biogenic Apatites: Implications for Paleoseawater Studies. Earth and Planetary Science Letters, 142(3/4): 425-437. https://doi.org/10.1016/0012-821x(96)00119-7 http://www.sciencedirect.com/science/article/pii/0012821X96001197 |
| Holser, W. T., 1997. Evaluation of the Application of Rare-Earth Elements to Paleoceanography. Palaeogeography, Palaeoclimatology, Palaeoecology, 132(1/2/3/4): 309-323. https://doi.org/10.1016/s0031-0182(97)00069-2 http://www.sciencedirect.com/science/article/pii/S0031018297000692 |
| Keenan, S. W., 2016. From Bone to Fossil: A Review of the Diagenesis of Bioapatite. American Mineralogist, 101(9): 1943-1951. https://doi.org/10.2138/am-2016-5737 |
| Keenan, S. W., Engel, A. S., 2017. Early Diagenesis and Recrystallization of Bone. Geochimica et Cosmochimica Acta, 196: 209-223. https://doi.org/10.1016/j.gca.2016.09.033 |
| Kim, J. -H., Torres, M. E., Haley, B. A., et al., 2012. The Effect of Diagenesis and Fluid Migration on Rare Earth Element Distribution in Pore Fluids of the Northern Cascadia Accretionary Margin. Chemical Geology, 291: 152-165. https://doi.org/10.1016/j.chemgeo.2011.10.010 |
| Knüpfer, J., 1967. Zur Fauna und Biostratigraphie des Ordoviziums (Gräfenthaler Schichten) in Thüringen. Freiberger Forschungshefte, C220: 1-119 |
| Kocsis, L., Trueman, C. N., Palmer, M. R., 2010. Protracted Diagenetic Alteration of REE Contents in Fossil Bioapatites: Direct Evidence from Lu-Hf Isotope Systematics. Geochimica et Cosmochimica Acta, 74(21): 6077-6092. https://doi.org/10.1016/j.gca.2010.08.007 |
| Kohn, M. J., Moses, R. J., 2013. Trace Element Diffusivities in Bone Rule out Simple Diffusive Uptake during Fossilization but Explain in vivo Uptake and Release. Proceedings of the National Academy of Sciences, 110(2): 419-424. https://doi.org/10.1073/pnas.1209513110 |
| Kowal-Linka, M., Jochum, K. P., Surmik, D., 2014. LA-ICP-MS Analysis of Rare Earth Elements in Marine Reptile Bones from the Middle Triassic Bonebed (Upper Silesia, S Poland): Impact of Long-Lasting Diagenesis, and Factors Controlling the Uptake. Chemical Geology, 363: 213-228. https://doi.org/10.1016/j.chemgeo.2013.10.038 |
| Lécuyer, C., Reynard, B., Grandjean, P., 2004. Rare Earth Element Evolution of Phanerozoic Seawater Recorded in Biogenic Apatites. Chemical Geology, 204(1/2): 63-102. https://doi.org/10.1016/j.chemgeo.2003.11.003 http://www.sciencedirect.com/science/article/pii/S0009254103003504 |
| LeGeros, R. Z., 1981. Apatites in Biological Systems. Progress in Crystal Growth and Characterization 4(1/2): 1-45 http://www.sciencedirect.com/science/article/pii/0146353581900460?_fmt=full |
| Li, Y., Zhao, L. S., Chen, Z. -Q., et al., 2017. Oceanic Environmental Changes on a Shallow Carbonate Platform (Yangou, Jiangxi Province, South China) during the Permian-Triassic Transition: Evidence from Rare Earth Elements in Conodont Bioapatite. Palaeogeography, Palaeoclimatology, Palaeoecology, 486: 6-16. https://doi.org/10.1016/j.palaeo.2017.02.035 |
| Liao, J. L., Sun, X. M., Li, D. F., et al., 2019. New Insights into Nanostructure and Geochemistry of Bioapatite in REE-Rich Deep-Sea Sediments: LA-ICP-MS, TEM, and Z-Contrast Imaging Studies. Chemical Geology, 512: 58-68. https://doi.org/10.1016/j.chemgeo.2019.02.039 |
| Lindström, M., Pelhate, A., 1971. Présence de Conodontes dans les Calcaires de Rosan (Ordovicien moyen a Supérieur, Massif Armoricain). Colloque Ordovicien-Silurien, Brest 1971. Mémoire du Bureau de Recherches Géologiques et Minières, 73: 89-91 |
| Malferrari, D., Ferretti, A., Mascia, M. T., et al., 2019. How much can We Trust Major Element Quantification in Bioapatite Investigation?. ACS Omega, 4(18): 17814-17822. https://doi.org/10.1021/acsomega.9b02426 |
| Martin, E. E., Scher, H. D., 2004. Preservation of Seawater Sr and Nd Isotopes in Fossil Fish Teeth: Bad News and Good News. Earth and Planetary Science Letters, 220(1/2): 25-39. https://doi.org/10.1016/s0012-821x(04)00030-5 http://www.sciencedirect.com/science/article/pii/S0012821X04000305 |
| McArthur, J. M., Walsh, J. N., 1984. Rare-Earth Geochemistry of Phosphorites. Chemical Geology, 47(3/4): 191-220. https://doi.org/10.1016/0009-2541(84)90126-8 http://www.sciencedirect.com/science/article/pii/0009254184901268 |
| McLennan, S. M., 2001. Relationships between the Trace Element Composition of Sedimentary Rocks and Upper Continental Crust. Geochemistry, Geophysics, Geosystems, 2(4). https://doi.org/10.1029/2000gc000109 doi: 10.1029/2000GC000109 |
| Medici, L., Malferrari, D., Savioli, M., et al., 2020. Mineralogy and Crystallization Patterns in Conodont Bioapatite from First Occurrence (Cambrian) to Extinction (end-Triassic). Palaeogeography, Palaeoclimatology, Palaeoecology, 549: 109098. https://doi.org/10.1016/j.palaeo.2019.02.024 |
| Nardelli, M. P., Malferrari, D., Ferretti, A., et al., 2016. Zinc Incorporation in the Miliolid Foraminifer Pseudotriloculina rotunda under Laboratory Conditions. Marine Micropaleontology, 126: 42-49. https://doi.org/10.1016/j.marmicro.2016.06.001 |
| Nothdurft, L. D., Webb, G. E., Kamber, B. S., 2004. Rare Earth Element Geochemistry of Late Devonian Reefal Carbonates, Canning Basin, Western Australia: Confirmation of a Seawater REE Proxy in Ancient Limestones. Geochimica et Cosmochimica Acta, 68(2): 263-283. https://doi.org/10.1016/s0016-7037(03)00422-8 doi: 10.1016/S0016-7037(03)00422-8 |
| Nozaki, Y., Zhang, J., Amakawa, H., 1997. The Fractionation between Y and Ho in the Marine Environment. Earth and Planetary Science Letters, 148(1/2): 329-340. https://doi.org/10.1016/s0012-821x(97)00034-4 http://www.sciencedirect.com/science/article/pii/S0012821X97000344 |
| Paris, F., Pelhate, A., Weyant, M., 1981. Conodontes ashgilliens dans la Formation de Rosan, coupe de Lostmarc'h (Finistère, Massif Armoricain). Conséquences Paléogéographiques. Bulletin de la Société Géologique et Mineralogique de Bretagne, 13(2): 15-35 http://www.researchgate.net/publication/286706487_Conodontes_ashgilliens_dans_la_Formation_de_Rosan_coupe_de_Lostmarc'h_Finistere_Massif_armoricain_Consequences_paleogeographiques |
| Pattan, J. N., Pearce, N. J. G., Mislankar, P. G., 2005. Constraints in Using Cerium-Anomaly of Bulk Sediments as an Indicator of Paleo Bottom Water Redox Environment: A Case Study from the Central Indian Ocean Basin. Chemical Geology, 221(3/4): 260-278. https://doi.org/10.1016/j.chemgeo.2005.06.009 http://www.sciencedirect.com/science/article/pii/S000925410500224X |
| Peppe, D. J., Reiners, P. W., 2007. Conodont (U-Th)/He Thermochronology: Initial Results, Potential, and Problems. Earth and Planetary Science Letters, 258(3/4): 569-580. https://doi.org/10.1016/j.epsl.2007.04.022 http://www.sciencedirect.com/science/article/pii/S0012821X07002373 |
| Person, A., Bocherens, H., Saliège, J. F., et al., 1995. Early Diagenetic Evolution of Bone Phosphate: An X-Ray Diffractometry Analysis. Journal of Archaeological Science, 22(2): 211-221. https://doi.org/10.1006/jasc.1995.0023 |
| Picard, S., Lécuyer, C., Barrat, J. A., et al., 2002. Rare Earth Element Contents of Jurassic Fish and Reptile Teeth and Their Potential Relation to Seawater Composition (Anglo-Paris Basin, France and England). Chemical Geology, 186(1/2): 1-16. https://doi.org/10.1016/s0009-2541(01)00424-7 http://www.sciencedirect.com/science/article/pii/S0009254101004247 |
| Pietsch, C., Bottjer, D. J., 2010. Comparison of Changes in Ocean Chemistry in the Early Triassic with Trends in Diversity and Ecology. Journal of Earth Science, 21(S1): 147-150. https://doi.org/10.1007/s12583-010-0195-9 |
| Pucéat, E., Reynard, B., Lécuyer, C., 2004. Can Crystallinity be Used to Determine the Degree of Chemical Alteration of Biogenic Apatites?. Chemical Geology, 205(1/2): 83-97. https://doi.org/10.1016/j.chemgeo.2003.12.014 http://www.sciencedirect.com/science/article/pii/S0009254103003991 |
| Reynard, B., Lécuyer, C., Grandjean, P., 1999. Crystal-Chemical Controls on Rare-Earth Element Concentrations in Fossil Biogenic Apatites and Implications for Paleoenvironmental Reconstructions. Chemical Geology, 155(3/4): 233-241. https://doi.org/10.1016/s0009-2541(98)00169-7 http://www.sciencedirect.com/science/article/pii/S0009254198001697 |
| Sanz-López, J., Blanco-Ferrera, S., 2012. Overgrowths of Large Authigenic Apatite Crystals on the Surface of Conodonts from Cantabrian Limestones (Spain). Facies, 58(4): 707-726. https://doi.org/10.1007/s10347-012-0295-3 |
| Shen, J., Algeo, T. J., Zhou, L., et al., 2012. Volcanic Perturbations of the Marine Environment in South China Preceding the Latest Permian Mass Extinction and Their Biotic Effects. Geobiology, 10(1): 82-103. https://doi.org/10.1111/j.1472-4669.2011.00306.x |
| Shields, G., Stille, P., 2001. Diagenetic Constraints on the Use of Cerium Anomalies as Palaeoseawater Redox Proxies: An Isotopic and REE Study of Cambrian Phosphorites. Chemical Geology, 175(1/2): 29-48. https://doi.org/10.1016/s0009-2541(00)00362-4 http://www.sciencedirect.com/science/article/pii/S0009254100003624 |
| Shields, G., Webb, G. E., 2004. Has the REE Composition of Seawater Changed over Geological Time?. Chemical Geology, 204: 103-107. https://doi.org/10.1016/j.chemgeo.2003.09.010 |
| Sholkovitz, E., Shen, G. T., 1995. The Incorporation of Rare Earth Elements in Modern Coral. Geochimica et Cosmochimica Acta, 59(13): 2749-2756. https://doi.org/10.1016/0016-7037(95)00170-5 |
| Smith, C. I., Craig, O. E., Prigodich, R. V., et al., 2005. Diagenesis and Survival of Osteocalcin in Archaeological Bone. Journal of Archaeological Science, 32(1): 105-113. https://doi.org/10.1016/j.jas.2004.07.003 |
| Song, H. J., Wignall, P. B., Song, H. Y., et al., 2019. Seawater Temperature and Dissolved Oxygen over the Past 500 Million Years. Journal of Earth Science, 30(2): 236-243. https://doi.org/10.1007/s12583-018-1002-2 |
| Sweet, W. C., Bergström, S. M., 1984. Conodont Provinces and Biofacies of the Late Ordovician. In: Clark, D. L., ed., Conodont Biofacies and Provincialism. Geological Society of America Special Paper, 196: 69-87 |
| Sweet, W. C., Donoghue, P. C. J., 2001. Conodonts: Past, Present, Future. Journal of Paleontology, 75(6): 1174-1184. https://doi.org/10.1017/s0022336000017224 doi: 10.1017/S0022336000017224 |
| Toyoda, K., Tokonami, M., 1990. Diffusion of Rare-Earth Elements in Fish Teeth from Deep-Sea Sediments. Nature, 345: 607-609. https://doi.org/10.1038/345607a0 |
| Trotter, J. A., Barnes, C. R., McCracken, A. D., 2016. Rare Earth Elements in Conodont Apatite: Seawater or Pore-Water Signatures?. Palaeogeography, Palaeoclimatology, Palaeoecology, 462: 92-100. https://doi.org/10.1016/j.palaeo.2016.09.007 |
| Trotter, J. A., Eggins, S. M., 2006. Chemical Systematics of Conodont Apatite Determined by Laser Ablation ICPMS. Chemical Geology, 233(3/4): 196-216. https://doi.org/10.1016/j.chemgeo.2006.03.004 http://www.sciencedirect.com/science/article/pii/S0009254106001501 |
| Trotter, J. A., Gerald, J. D. F., Kokkonen, H., et al., 2007. New Insights into the Ultrastructure, Permeability, and Integrity of Conodont Apatite Determined by Transmission Electron Microscopy. Lethaia, 40(2): 97-110. https://doi.org/10.1111/j.1502-3931.2007.00024.x |
| Trueman, C. N., Privat, K., Field, J., 2008. Why do Crystallinity Values Fail to Predict the Extent of Diagenetic Alteration of Bone Mineral? Palaeogeography, Palaeoclimatology, Palaeoecology, 266(3/4): 160-167. https://doi.org/10.1016/j.palaeo.2008.03.038 http://new.med.wanfangdata.com.cn/Paper/Detail?id=PeriodicalPaper_JJ025615518 |
| Trueman, C. N., Tuross, N., 2002. Trace Elements in Recent and Fossil Bone. In: Kohn, M. J., Rakovan, J., Hughes, J. M., eds., Phosphates: Geochemical, Geobiological and Materials Importance. Review in Mineralogy and Geochemistry, 48: 489-521 |
|
Vidal, M., Dabard, M. -P., Gourvennec, R., et al., 2011. Le |
| Webb, G. E., Kamber, B. S., 2000. Rare Earth Elements in Holocene Reefal Microbialites: A New Shallow Seawater Proxy. Geochimica et Cosmochimica Acta, 64(9): 1557-1565. https://doi.org/10.1016/s0016-7037(99)00400-7 doi: 10.1016/S0016-7037(99)00400-7 |
| Webb, G. E., Nothdurft, L. D., Kamber, B. S., et al., 2009. Rare Earth Element Geochemistry of Scleractinian Coral Skeleton during Meteoric Diagenesis: A Sequence through Neomorphism of Aragonite to Calcite. Sedimentology, 56(5): 1433-1463. https://doi.org/10.1111/j.1365-3091.2008.01041.x |
| Wenzel, B., Lécuyer, C., Joachimski, M. M., 2000. Comparing Oxygen Isotope Records of Silurian Calcite and Phosphate—δ18O Compositions of Brachiopods and Conodonts. Geochimica et Cosmochimica Acta, 64(11): 1859-1872. https://doi.org/10.1016/s0016-7037(00)00337-9 doi: 10.1016/S0016-7037(00)00337-9 |
| Weyant, M., Dorè, F., Le Gall, J., et al., 1977. Un épisode Calcaire ashgillien dans l'est du Massif Armoricain: Incidence Sur l'ậge des Dépôts Glacio-Marins fini-Ordoviciens. Comptes Rendus de l'Académie des Sciences, 284(D): 1147-1149 http://www.researchgate.net/publication/284504245_Un_episode_calcaire_ashgillien_dans_l'est_du_Massif_armoricain_incidence_sur_l'age_des_depots_glacio-marins_fini-ordoviciens |
| Wheeley, J. R., Smith, M. P., Boomer, I., 2012. Oxygen Isotope Variability in Conodonts: Implications for Reconstructing Palaeozoic Palaeoclimates and Palaeoceanography. Journal of the Geological Society, 169(3): 239-250. https://doi.org/10.1144/0016-76492011-048 |
| Wright, J., Schrader, H., Holser, W. T., 1987. Paleoredox Variations in Ancient Oceans Recorded by Rare Earth Elements in Fossil Apatite. Geochimica et Cosmochimica Acta, 51(3): 631-644. https://doi.org/10.1016/0016-7037(87)90075-5 |
| Wright, J., Colling, A., 1995. Seawater: Its Composition, Properties and Behavior. Second ed. Pergamon, Oxford. 168 |
| Wright, J., Seymour, R. S., Shaw, H. F., 1984. REE and Nd Isotopes in Conodont Apatite: Variations with Geological Age and Depositional Environment. In: Clark, D. L., ed., Conodont Biofacies and Provincialism. Geological Society of America Special Paper, 196: 325-340 |
| Xin, H., Jiang, S. Y., Yang, J. H., et al., 2016. Rare Earth Element Geochemistry of Phosphatic Rocks in Neoproterozoic Ediacaran Doushantuo Formation in Hushan Section from the Yangtze Gorges Area, South China. Journal of Earth Science, 27(2): 204-210. https://doi.org/10.1007/s12583-015-0653-5 |
| Zhang, L., Algeo, T. J., Cao, L., et al., 2016. Diagenetic Uptake of Rare Earth Elements by Conodont Apatite. Palaeogeography, Palaeoclimatology, Palaeoecology, 458:176-197. https://doi.org/10.1016/j.palaeo.2015.10.049 |
| Zhang, L., Cao, L., Zhao, L. S., et al., 2017. Raman Spectral, Elemental, Crystallinity, and Oxygen-Isotope Variations in Conodont Apatite during Diagenesis. Geochimica et Cosmochimica Acta, 210:184-207. https://doi.org/10.1016/j.gca.2017.04.036 |
| Zhang, J., Nozaki, Y., 1996. Rare Earth Elements and Yttrium in Seawater: ICP-MS Determinations in the East Caroline, Coral Sea, and South Fiji Basins of the Western South Pacific Ocean. Geochimica et Cosmochimica Acta, 60(23): 4631-4644. https://doi.org/10.1016/s0016-7037(96)00276-1 doi: 10.1016/S0016-7037(96)00276-1 |
| Zhang, J., Amakawa, H., Nozaki, Y., 1994. The Comparative Behaviors of Yttrium and Lanthanides in the Seawater of the North Pacific. Geophysical Research Letters, 21(24): 2677-2680. https://doi.org/10.1029/94gl02404 doi: 10.1029/94GL02404 |
| Zhao, L. S., Chen, Z.-Q., Algeo, T. J., et al., 2013. Rare-Earth Element Patterns in Conodont Albid Crowns: Evidence for Massive Inputs of Volcanic Ash during the Latest Permian Biocrisis?. Global and Planetary Change, 105:135-151. https://doi.org/10.1016/j.gloplacha.2012.09.001 |
| Žigaitė, Ž., Qvarnström, M., Bancroft, A., et al., 2020. Trace and Rare Earth Element Compositions of Silurian Conodonts from the Vesiku Bone Bed: Histological and Palaeoenvironmental Implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 549:109449. https://doi.org/10.1016/j.palaeo.2019.109449 |
Figures(5) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: