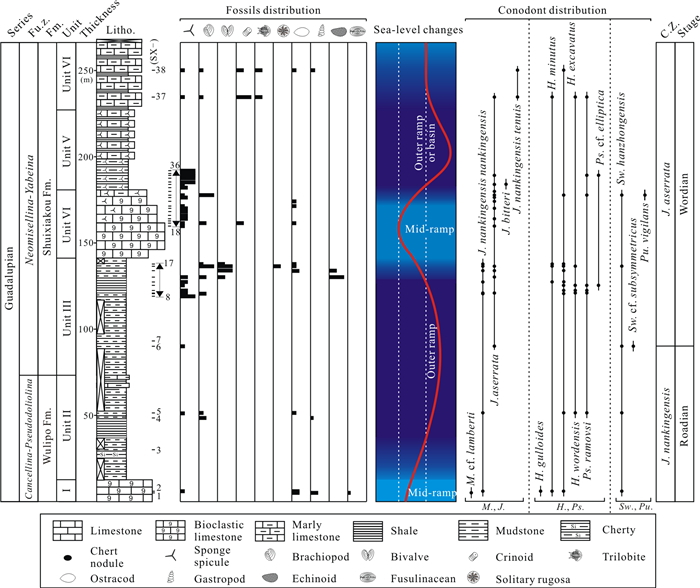
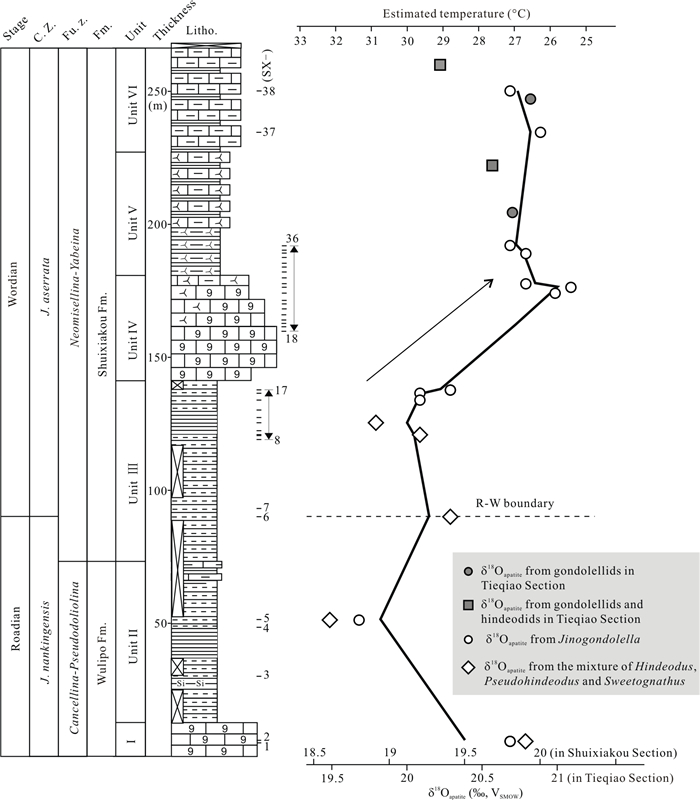
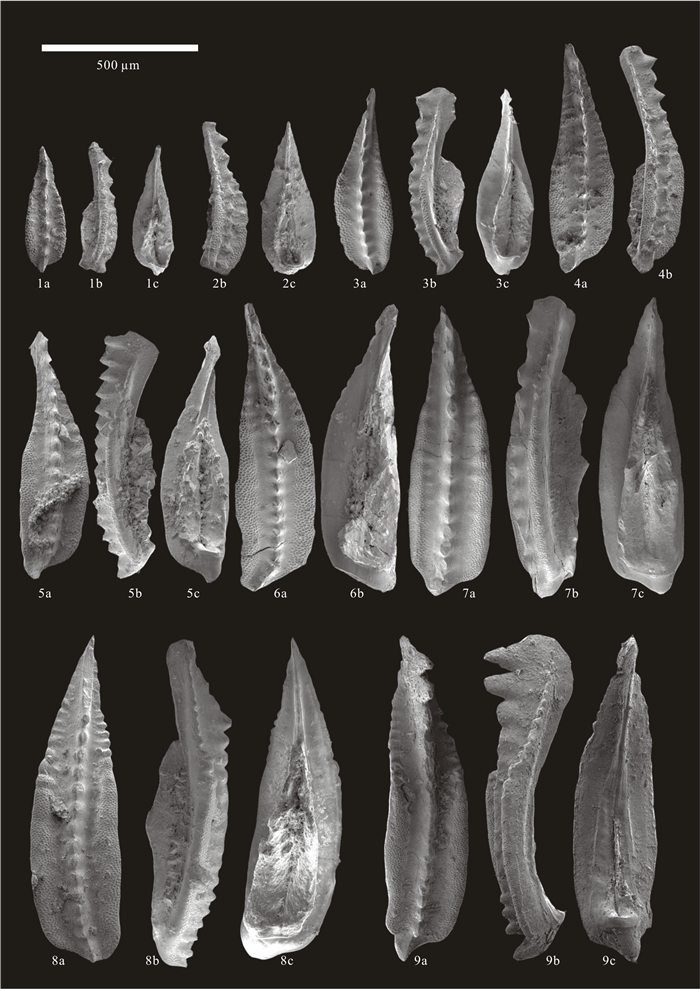
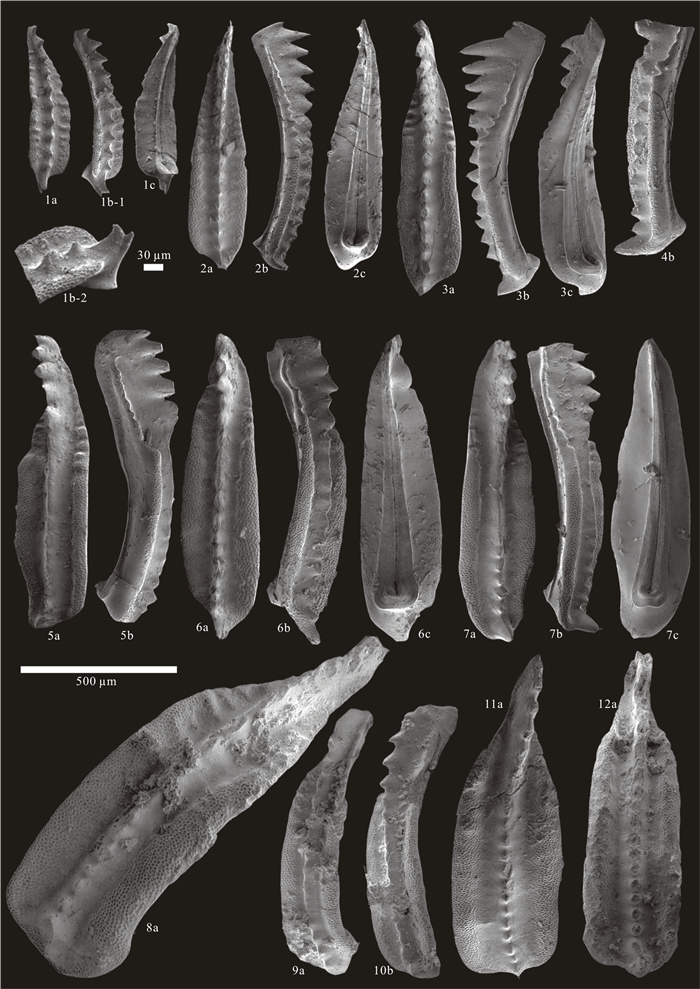
| Citation: | Baojin Wu, Hanxiao Li, Michael M. Joachimski, Paul B. Wignall, Haishui Jiang, Jiaxin Yan, Lina Wang, Xianlang Wu, Xulong Lai. Roadian-Wordian (Middle Permian) Conodont Biostratigraphy, Sedimentary Facies and Paleotemperature Evolution at the Shuixiakou Section, Xikou Area, Southeastern Qinling Region, China. Journal of Earth Science, 2021, 32(3): 534-553. doi: 10.1007/s12583-020-1099-y

|

| Angiolini, L., Zanchi, A., Zanchetta, S., et al., 2015. From Rift to Drift in South Pamir (Tajikistan): Permian Evolution of a Cimmerian Terrane. Journal of Asian Earth Sciences, 102: 146-169. https://doi.org/10.1016/j.jseaes.2014.08.001 |
| Anonymous, 1965. The 1: 200 000 Geological Map of Ankang Sheet. Regional Bureau of Ministry of Geology in Geological Survey, Northwest Institute of Geological Sciences (in Chinese) |
| Behnken, F. H., 1975. Leonardian and Guadalupian (Permian) Conodont Biostratigraphy in Western and Southwestern United States. Journal of Paleontology, 49(2): 284-315. https://doi.org/10.2307/1303362 |
| Behnken, F. H., Wardlaw, B. R., Stout, L. N., 1986. Conodont Biostratigraphy of the Permian Meade Peak Phosphatic Shale Member, Phosphoria Formation, Southeastern Idaho. Rocky Mountain Geology, 24(2): 169-190 http://rmg.geoscienceworld.org/content/24/2/169 |
| Biakov, A. S., 2015. Biogeography of the Permian Marine Boreal Basins Based on Bivalves. Paleontological Journal, 49(11): 1184-1192. https://doi.org/10.1134/s0031030115110040 |
| Burrett, C., Udchachon, M., Thassanapak, H., et al., 2014. Conodonts, Radiolarians and Ostracodes in the Permian E-Lert Formation, Loei Fold Belt, Indochina Terrane, Thailand. Geological Magazine, 152(1): 106-142. https://doi.org/10.1017/s001675681400017x |
| Chen, Z. -Q., George, A. D., Yang, W. R., 2009. Effects of Middle-Late Permian Sea-Level Changes and Mass Extinction on the Formation of the Tieqiao Skeletal Mound in the Laibin Area, South China. Australian Journal of Earth Sciences, 56(6): 745-763. https://doi.org/10.1080/08120090903002581 |
| Chen, Z. -Q., Jin, Y. G., Shi, G. -R., 1998. Permian Transgression-Regression Sequences and Sea-Level Changes of South China. Proceedings of the Royal Society of Victoria, 110: 345-367 http://www.researchgate.net/publication/290798729_Permian_transgression-regression_sequences_and_sea-level_changes_of_South_China |
| Chen, B., Joachimski, M. M., Shen, S. Z., et al., 2013. Permian Ice Volume and Palaeoclimate History: Oxygen Isotope Proxies Revisited. Gondwana Research, 24(1): 77-89. https://doi.org/10.1016/j.gr.2012.07.007 |
| Chen, B., Joachimski, M. M., Sun, Y. D., et al., 2011. Carbon and Conodont Apatite Oxygen Isotope Records of Guadalupian-Lopingian Boundary Sections: Climatic or Sea-Level Signal?. Palaeogeography, Palaeoclimatology, Palaeoecology, 311(3/4): 145-153. https://doi.org/10.1016/j.palaeo.2011.08.016 |
| Cheng, C., Li, S. Y., Cao, T. L., 2017. Permian Conodonts at the Xikou Section, Zhen'an County, Shananxi Province, NW China. Acta Micropalaeontologica Sinica, 34(1): 16-32 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-WSGT201701002.htm |
| Cheng, C., Li, S. Y., Xie, X. Y., et al., 2019. Permian Carbon Isotope and Clay Mineral Records from the Xikou Section, Zhenʼan, Shaanxi Province, Central China: Climatological Implications for the Easternmost Paleo-Tethys. Palaeogeography, Palaeoclimatology, Palaeoecology, 514: 407-422. https://doi.org/10.1016/j.palaeo.2018.10.023 |
| Chernykh, V. V., Chuvashov, B. I., Davydov, V. I., et al., 2006. Usolka Section (southern Urals, Russia): A Potential Candidate for GSSP to Define the Base of the Gzhelian Stage in the Global Chronostratigraphic Scale. Geologija, 49(2): 205-217. https://doi.org/10.5474/geologija.2006.015 |
| Ching, Y. K., 1960. Conodonts from the Kufeng Suite (Formation) of Lungtan, Nanking. Acta Palaeontologica Sinica, 8: 242-248 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTotal-GSWX196003002.htm |
| Clark, D. L., 1972. Early Permian Crisis and Its Bearing on Permo-Triassic Conodont Taxonomy. Geologica et Palaeontòlogica, l: 147-158 http://www.researchgate.net/publication/281603830_Early_Permian_crisis_and_its_bearing_on_Permo-Triassic_conodont_taxonomy |
| Clark, D. L., Behnken, F. H., 1979. Evolution and Taxonomy of the North USA Upper Permian Neogondolella Serrata Complex. Journal of Paleontology, 53(2): 263-275 |
| Clark, D. L., Ethington, R. L., 1962. Survey of Permian Conodonts in Western North America. Brigham Young University Geology Studies, 9(2): 102-114 http://www.researchgate.net/publication/312614490_Survey_of_Permian_conodonts_in_western_North_America |
| Clark, D. L., Mosher, L. C., 1966. Stratigraphic, Geographic, and Evolutionary Development of the Conodont Genus Gondolella. Journal of Paleontology, 40(2): 376-394 |
| Clark, D. L., Wang, C. Y., 1988. Permian Neogondolellids from South China: Significance for Evolution of the Serrata and Carinata Groups in North America. Journal of Paleontology, 62(1): 132-138. https://doi.org/10.1017/s0022336000058972 |
| Davydov, V. I., Biakov, A. S., Schmitz, M. D., et al., 2018. Radioisotopic Calibration of the Guadalupian (Middle Permian) Series: Review and Updates. Earth-Science Reviews, 176: 222-240. https://doi.org/10.1016/j.earscirev.2017.10.011 |
| Ding, P. Z., Jin, T. A., Sun, X. F., 1983. The Marine Permian of Zhenan, Southern Shaanxi, Eastern Qinling Range. Bulletin of Xi'an Institute of Geology and Mineral Resources, Chinese Academy of Geological Sciences, (6): 99-104 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTotal-XBFK198306008.htm |
| Ding, P. Z., Jin, T. A., Sun, X. F., 1987. An Excursion Guide to Permian Geology of Xikou Area, Zhen'an County, Shananxi. Bulletin of Xi'an Institute of Geology and Mineral Resources, Chinese Academy of Geological Sciences, 15: 113-138 (in Chinese with English Abstract) http://ci.nii.ac.jp/naid/10006848287 |
| Ding, P. Z., Jin, T. A., Sun, X. F., 1989. The Marine Permian Strata and Its Faunal Assemblages in Xikou Area of Zhen'an County, South Shaanxi, East Qinling range. Bulletin of Xi'an Institute of Geology and Mineral Resources, Chinese Academy of Geological Sciences, (25): 1-68 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-XBFK198901000.htm |
| Dong, Y. P., Santosh, M., 2016. Tectonic Architecture and Multiple Orogeny of the Qinling Orogenic Belt, Central China. Gondwana Research, 29(1): 1-40. https://doi.org/10.1016/j.gr.2015.06.009 |
| Dzki, J., 1976. Remarks on the Evolution of Ordovician Conodonts. Acta Palaeontologica Polonica, 21: 395-455 http://ci.nii.ac.jp/naid/10019474740 |
| Eichenberg, W., 1930. Conodonten Aus Dem Culm des Harzes. Paläontologische Zeitschrift, 12(3/4): 177-182. https://doi.org/10.1007/bf03044446 |
| Ellison, S., 1941. Revision of the Pennsylvanian Conodonts. Journal of Paleontology, 15(2): 107-143. https://doi.org/10.2307/1298938 |
| Fielding, C. R., Frank, T. D., Birgenheier, L. P., et al., 2008. Stratigraphic Imprint of the Late Palaeozoic Ice Age in Eastern Australia: A Record of Alternating Glacial and Nonglacial Climate Regime. Journal of the Geological Society, 165(1): 129-140. https://doi.org/10.1144/0016-76492007-036 |
| Frank, T. D., Shultis, A. I., Fielding, C. R., 2015. Acme and Demise of the Late Palaeozoic Ice Age: A View from the Southeastern Margin of Gondwana. Palaeogeography, Palaeoclimatology, Palaeoecology, 418: 176-192. https://doi.org/10.1016/j.palaeo.2014.11.016 |
| Glenister, B. F., Wardlaw, B. R., Lambert, L. L., et al., 1999. Proposal of Guadalupian and Component Roadian, Wordian, and Capitanian Stages as International Standards for the Middle Permian Series. Permophiles, 34: 3-11 http://www.researchgate.net/publication/285294760_Proposal_of_Guadalupian_and_component_Roadian_Wordian_and_Capitanian_stages_as_international_standards_for_the_Middle_Permian_Series |
| Grossman, E. L., Yancey, T. E., Jones, T. E., et al., 2008. Glaciation, Aridification, and Carbon Sequestration in the Permo-Carboniferous: The Isotopic Record from Low Latitudes. Palaeogeography, Palaeoclimatology, Palaeoecology, 268(3/4): 222-233. https://doi.org/10.1016/j.palaeo.2008.03.053 |
| Golding, M. L., 2018. The Multielement Apparatuses of Guadalupian to Lopingian (Middle-Upper Permian) Sweetognathids from North America, and Their Significance for the Phylogeny of Late Paleozoic Conodonts. Bulletins of American Paleontology, 395/396: 115-125. https://doi.org/10.32857/bap.2018.395.09 |
| Gullo, M., Kozur, H., 1992. Conodonts from the Pelagic Deep-Water Permian of Central Western Sicily (Italy). Neues Jahrbuchfure Geologie und Palaeontologie, 184(2): 203-234 http://www.researchgate.net/publication/284790280_Conodonts_from_the_pelagic_deep-water_Permian_of_central_Western_Sicily_Italy |
| Henderson, C. M., 1981. Conodont Paleontology of the Permian Sabine Bay, Assistance and Trold Fiord Formations, Northern Ellesmere Island, Canadian Arctic Archipelago: [Dissertation]. University of British Columbia, Vancouver |
| Henderson, C. M., 2018. Permian Conodont Biostratigraphy. Geological Society, London, Special Publications, 450: 119-142. https://doi.org/10.1144/sp450.9 |
| Henderson, C. M., Mei, S. L., 2003. Stratigraphic versus Environmental Significance of Permian Serrated Conodonts around the Cisuralian-Guadalupian Boundary: New Evidence from Oman. Palaeogeography, Palaeoclimatology, Palaeoecology, 191(3/4): 301-328. https://doi.org/10.1016/s0031-0182(02)00669-7 |
| Henderson, C. M., Mei, S. L., 2007. Geographical Clines in Permian and Lower Triassic Gondolellids and Its Role in Taxonomy. Palaeoworld, 16(1/2/3): 190-201. https://doi.org/10.1016/j.palwor.2007.05.014 |
| Isakova, T. N., 1998. Review of the Conodonts of the Sakmarian Stratotype Section (South Urals). Palaeontologia Polonica, 58: 261-271 http://www.palaeontologia.pan.pl/Archive/1998-58_261-271.pdf |
| Joachimski, M. M., Breisig, S., Buggisch, W., et al., 2009. Devonian Climate and Reef Evolution: Insights from Oxygen Isotopes in Apatite. Earth and Planetary Science Letters, 284(3/4): 599-609. https://doi.org/10.1016/j.epsl.2009.05.028 |
| Kozur, H. W., 1975. Beitrage zur Conodont en Fauna des Perm. Geologische Palaontologische Mitteilungen Innsbruck, 5: 1-44 http://ci.nii.ac.jp/naid/10006849341 |
| Kozur, H. W., 1992. Dzhulfian and Early Changxingian (Late Permian) Tethyan Conodonts from the Glass Mountains, West Taxas. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 187(1): 99-114 http://ci.nii.ac.jp/naid/80007287021 |
| Kozur, H. W., 1995. Permian Conodont Zonation and Its Importance for the Permian Stratigraphic Standard Scale. Geologische Palaontologische Mitteilungen Innsbruck, 20: 165-205 http://www.researchgate.net/publication/265306630_Permian_conodont_zonation_and_its_importance_for_the_Permian_stratigraphic_standard_scale |
| Kozur, H. W., Mostler, H., 1995. Guadalupian (Middle Permian) Conodonts of Sponge-Bearing Limestones from the Margins of the Delaware Basin, West Texas. Geologia Croatica, 48(2): 107-128 |
| Kozur, H. W., Wardlaw, B. R., 2010. The Guadalupian Conodont Fauna of Rustaq and Wadi Wasit, Oman and a West Texas Connection. Micropaleontology, 56(1/2): 213-231. https://doi.org/10.2307/40607082 |
| Kutygin, R. V., 2015. Biogeographic Distribution and Relationship of Permian Ammonoid Communities of Verkhoyano-Okhotsk and Kolymo-Omolon Regions. Science Education, (2): 46-50 (in Russian) |
| Lai, X. L., Wang, W., Wignall, P. B., et al., 2008. Palaeoenvironmental Change during the End-Guadalupian (Permian) Mass Extinction in Sichuan, China. Palaeogeography, Palaeoclimatology, Palaeoecology, 269(1/2): 78-93. https://doi.org/10.1016/j.palaeo.2008.08.005 |
| Lambert, L. L., Bell, G. L. Jr, Fronimos, J. A., et al., 2010. Conodont Biostratigraphy of a more Complete Reef Trail Member Section near the Type Section, Latest Guadalupian Series Type Region. Micropaleontology, 56: 233-253. https://doi.org/10.2307/40607083 |
| Lambert, L. L., Wardlaw, B. R., Henderson, C. M., 2007. Mesogondolella and Jinogondolella (Conodonta): Multielement Definition of the Taxa that Bracket the Basal Guadalupian (Middle Permian Series) GSSP. Palaeoworld, 16(1/2/3): 208-221. https://doi.org/10.1016/j.palwor.2007.05.017 |
| Lambert, L. L., Wardlaw, B. R., Nestell, M. K., et al., 2002. Latest Guadalupian (Middle Permian) Conodonts and Foraminifers from West Texas. Micropaleontology, 48(4): 343-364. https://doi.org/10.1661/0026-2803(2002)048[0343:lgmpca]2.0.co;2 |
| Lindström, M., 1970. A Suprageneric Taxonomy of the Conodonts. Lethaia, 3(4): 427-445. https://doi.org/10.1111/j.1502-3931.1970.tb00834.x |
| Ma, Q. F., Feng, Q. L., Caridroit, M., et al., 2016. Integrated Radiolarian and Conodont Biostratigraphy of the Middle Permian Gufeng Formation (South China). Comptes Rendus Palevol, 15(5): 453-459. https://doi.org/10.1016/j.crpv.2016.02.001 |
| Mei, S. L., Henderson, C. M., 2001. Evolution of Permian Conodont Provincialism and Its Significance in Global Correlation and Paleoclimate Implication. Palaeogeography, Palaeoclimatology, Palaeoecology, 170(3/4): 237-260. https://doi.org/10.1016/s0031-0182(01)00258-9 |
| Mei, S. L., Henderson, C. M., 2002. Conodont Definition of the Kungurian (Cisuralian) and Roadian (Guadalupian) Boundary. Canadian Society of Petroleum Geologists, Memoir, 19: 529-551 http://www.researchgate.net/publication/303722461_Conodont_definition_of_the_Kungurian_Cisuralian_and_Roadian_Guadalupian_boundary |
| Mei, S. L., Wardlaw, B. R., 1994. Jinogondolella: A New Genus of Permian Gondolellids. International Symposium on Permian Stratigraphy, Environments and Resources, Abstracts, Guiyang, China. 20-21 |
| Mei, S. L., Henderson, C. M., Jin, Y. G., 1999. Permian Conodont Provincialism, Zonation and Global Correlation. Permophiles, 35: 16 |
| Mei, S. L., Henderson, C. M., Wardlaw, B. R., 2002. Evolution and Distribution of the Conodonts Sweetognathus and Iranognathus and Related Genera during the Permian, and Their Implications for Climate Change. Palaeogeography, Palaeoclimatology, Palaeoecology, 180(1/2/3): 57-91. https://doi.org/10.1016/s0031-0182(01)00423-0 |
| Mei, S. L., Jin, Y. G., Wardlaw, R. B., 1994. Succession of Conodont Zones from the Permian "Kuhfeng" Formation, Xuanhan, Sichuan and Its Implication in Global Correlation. Acta Palaeontologica Sinica, 33(1): 1-23 (in Chinese with English Abstract) http://europepmc.org/abstract/cba/274412 |
| Mei, S. L., Jin, Y. G., Wardlaw, B. R., 1998. Conodont Succession of the Guadalupian-Lopingian Boundary Strata in Labin of Guangxi, China and West Texas, USA. Palaeoworld, 9: 53-56 http://ci.nii.ac.jp/naid/10020145826 |
| Metcalfe, I., 1996. Gondwanaland Dispersion, Asian Accretion and Evolution of Eastern Tethys*. Australian Journal of Earth Sciences, 43(6): 605-623. https://doi.org/10.1080/08120099608728282 |
| Metcalfe, I., 2013. Gondwana Dispersion and Asian Accretion: Tectonic and Palaeogeographic Evolution of Eastern Tethys. Journal of Asian Earth Sciences, 66: 1-33. https://doi.org/10.1016/j.jseaes.2012.12.020 |
| Mory, A. J., Redfern, J., Martin, J. R., et al., 2008. A Review of Permian-Carboniferous Glacial Deposits in Western Australia. Geological Society of America Bulletin, 441: 29-40. https://doi.org/10.1130/2008.2441(02) |
| Muttoni, G., Gaetani, M., Kent, D. V., et al., 2009. Opening of the Neo-Tethys Ocean and the Pangea B to Pangea A Transformation during the Permian. GeoArabia, 14(4): 17-48 http://www.researchgate.net/publication/228515990_Opening_of_the_Neo-Tethys_Ocean_and_the_Pangea_B_to_Pangea_A_transformation_during_the_Permian |
| Nakrem, H. A., 1991. Conodonts from the Permian Succession of Bjornoya (Svalbard). Norsk Geologisk Tidsskr, 71: 235-248 http://www.researchgate.net/publication/281509015_Distribution_of_conodonts_through_the_Permian_succession_of_Svalbard |
| Nicora, A., Baud, A., Henderson, C. M., et al., 2009. Distribution of Hindeoduswordensis Wardlaw, 2000 in Space and Time. ICOS2009 Abstracts, 1: 36 |
| Nishikane, Y., Kaiho, K., Henderson, C. M., et al., 2014. Guadalupian-Lopingian Conodont and Carbon Isotope Stratigraphies of a Deep Chert Sequence in Japan. Palaeogeography, Palaeoclimatology, Palaeoecology, 403: 16-29. https://doi.org/10.1016/j.palaeo.2014.02.033 |
| Orchard, M. J., 1987. Conodonts from Western Canadian Chert: Their Nature, Distribution and Stratigraphic Application. In: Austin, R. L., ed., Conodonts: Investigative Techniques and Applications. British Micropaleontological Society, Ellis Horwood, Chichester. 94-119 |
| Orchard, M. J., Cordey, F., Rui, L., et al., 2001. Biostratigraphic and Biogeographic Constraints on the Carboniferous to Jurassic Cache Creek Terrane in Central British Columbia. Canadian Journal of Earth Sciences, 38(4): 551-578. https://doi.org/10.1139/e00-120 |
| Pucéat, E., Joachimski, M. M., Bouilloux, A., et al., 2010. Revised Phosphate-Water Fractionation Equation Reassessing Paleotemperatures Derived from Biogenic Apatite. Earth and Planetary Science Letters, 298(1/2): 135-142. https://doi.org/10.1016/j.epsl.2010.07.034 |
| Rexroad, C. B., Furnish, W. M., 1964. Conodonts from the Pella Formation (Mississippian), South-Central Iowa. Journal of Paleontology, 38: 667-676. https://doi.org/10.2307/1301445 |
| Rhodes, F. H. T., 1963. Conodonts from the Topmost Tensleep Sandstone of the Eastern Big Horn Mountains, Wyoming. Journal of Paleontology, 401-408. https://doi.org/10.2307/1301302 |
| Ritter, S. M., 1986. Taxonomic Revision and Phylogeny of Post-Early Permian Crisis Bisselli-Whitei Zone Conodonts with Comments on Late Paleozoic Diversity. Geologica et Palaeontologica, (20): 139-165 http://ci.nii.ac.jp/naid/10012507031 |
| Rybalka, S. V., 1989. Conodonts and Some Problems of Paleozoic Stratigraphy of Primorye. Bulletin of Russian Academy of Natural Sciences, 64(2): 95-99 (in Russian) |
| Sano, H., Orchard, M. J., 2004. Necoslie Breccia: Mixed Conodont Fauna-Bearing Neptunian Dyke in Carboniferous-Permian Seamount-Capping Oceanic Buildup (Pope Succession, Cache Creek Complex, Central British Columbia). Facies, 50(1): 133-145. https://doi.org/10.1007/s10347-004-0002-0 |
| Shen, S. Z., Zhang, H., Zhang, Y. C., et al., 2018. Permian Integrative Stratigraphy and Timescale of China. Science China Earth Sciences, 62(1): 154-188. https://doi.org/10.1007/s11430-017-9228-4 |
| Stampfli, G. M., Hochard, C., Vérard, C., et al., 2013. The Formation of Pangea. Tectonophysics, 593(3): 1-19. https://doi.org/10.1016/j.tecto.2013.02.037 |
| Sun, Y. D., Lai, X. L., Jiang, H. S., et al., 2008. Guadalupian (Middle Permian) Conodont Faunas at Shangsi Section, Northeast Sichuan Province. Journal of China University of Geosciences, 19(5): 451-460. https://doi.org/10.1016/s1002-0705(08)60050-3 |
| Sun, Y. D., Lai, X. L., Wignall, P. B., et al., 2010. Dating the Onset and Nature of the Middle Permian Emeishan Large Igneous Province Eruptions in SW China Using Conodont Biostratigraphy and Its Bearing on Mantle Plume Uplift Models. Lithos, 119(1/2): 20-33. https://doi.org/10.1016/j.lithos.2010.05.012 |
| Sun, Y. D., Liu, X. T., Yan, J. X., et al., 2017. Permian (Artinskian to Wuchapingian) Conodont Biostratigraphy in the Tieqiao Section, Laibin Area, South China. Palaeogeography, Palaeoclimatology, Palaeoecology, 465: 42-63. https://doi.org/10.1016/j.palaeo.2016.10.013 |
| Sungatullina, G. M., Davydov, V. I., 2015. New Data on Conodonts from the Kasimovian Stage of the Usolka Section, Southern Ural Mountains. Paleontological Journal, 49(10): 1142-1149. https://doi.org/10.1134/s0031030115110106 |
| Wardlaw, B. R., 2000. Guadalupian Conodont Biostratigraphy of the Glass and Del Norte Mountains. Guadalupian Symposium, 32: 37-88. https://doi.org/10.5479/si.00810274.32.1 |
| Wardlaw, B. R., 2015. Gondolellid Conodonts and Depositional Setting of the Phosphoria Formation. Micropaleontology, 61(4/5): 335-368 |
| Wardlaw, B. R., Collinson, J. W., 1979. Youngest Permian Conodont Faunas from the Great Basin and Rocky Mountain Regions. Brigham Young University Geology Studies, 26: 151-163 http://www.researchgate.net/publication/313516822_Youngest_Permian_conodont_faunas_from_the_Great_Basin_and_Rocky_Mountain_regions |
| Wardlaw, B. R., Collinson, J. W., 1984. Conodont Paleoecology of the Permian Phosphoria Formation and Related Rocks of Wyoming and Adjacent Areas. Geological Society of America Special Paper, 196: 263-281 http://www.researchgate.net/publication/296061483_Conodont_paleoecology_of_the_Permian_Phosphoria_Formation_and_related_rocks_of_Wyoming_and_adjacent_areas |
| Wardlaw, B. R., Collinson, J. W., 1986. Paleontology and Deposition of the Phosphoria Formation. Rocky Mountain Geology, 24(2): 107-142 http://www.researchgate.net/publication/259842943_Paleontology_and_deposition_of_the_Phosphoria_Formation |
| Wardlaw, B. R., Nestell, M. K., 2010. Three Jinogondolella Apparatuses from a Single Bed of the Bell Canyon Formation in the Apache Mountains, West Texas. Micropaleontology, 56(1/2): 195-212. https://doi.org/10.2307/40607081 |
| Wardlaw, B. R., Nestell, M. K., 2015. Conodont Faunas from a Complete Basinal Succession of the Upper Part of the Wordian (Middle Permian, Guadalupian, West Texas). Micropaleontology, 61: 257-292 http://smartsearch.nstl.gov.cn/paper_detail.html?id=ed85ced484e671199f4b3285fa742fff |
| Wardlaw, B. R., Mei, S. L., 1998. A Discussion of the Early Reported Species of Clarkina (Permian Conodonta) and the Possible Origin of the Genus. Palaeoworld, 9: 33-52 |
| Wang, C. Y., 1995. A Conodont Fauna from the Lowermost Kufeng Formation (Permian). Acta Micropalaeontologica Sinica, 12(3): 293-298 (in Chinese with English Abstract) http://search.cnki.net/down/default.aspx?filename=WSGT503.006&dbcode=CJFD&year=1995&dflag=pdfdown |
| Wang, C. Y., Ariunchimeg, Y. A., 2006. An Important Permian Conodont Species from Mongolia. Acta Micropalaeontologica Sinica, 23(3): 313-316 (in Chinese with English Abstract) http://manuscriptpro.com/profile/journal2/Wei-ti-gu-sheng-wu-xue-bao-=-Acta-micropalaeontologica-Sinica |
| Wang, C. Y., Dong, Z. C., 1991. Permian Conodonts from Suoxiyu in Cili County, Hunan. Acta Micropalaeontologica Sinica, 8(1): 41-58 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-WSGT199101003.htm |
| Wang, C. Y., Ritter, S. M., Clark, D. L., 1987. The Sweetognathus Complex in the Permian of China: Implications for Evolution and Homeomorphy. Journal of Paleontology, 61(5): 1047-1057. https://doi.org/10.1017/s0022336000029395 |
| Wang, C. Y., Wang, P., Li, W. G., 2004. Conodonts from the Permian Jisu Honguer (Zhesi) Formation of Inner Mongolia, China. Geobios, 37(4): 471-480. https://doi.org/10.1016/j.geobios.2003.06.003 |
| Wang, C. Y., Wu, J., Zhu, T., 1998. Permian Conodonts from the Penglaitan Section, Laipin County, Guangxi and the Base of the Wuchiapingian Stage (Lopingian Series). Acta Micropalaeontologica Sinica, 15(3): 225-235 (in Chinese with English Abstract) http://europepmc.org/abstract/cba/317991 |
| Wang, C. Y., Zhen, C. Z., Pen, Y. J., et al., 2000. A Conodont Fauna of Permian Northern Temperate Zone from the Fanjiatun Formation at Lijiayao, Jilin. Acta Micropalaeontologica Sinica, 17(4): 430-442 (in Chinese with English Abstract) http://www.researchgate.net/publication/286005524_A_conodont_fauna_of_Permian_Northern_temperate_Zone_from_the_Fanjiatun_Formation_at_Lijiayao_Jilin |
| Wang, D. C., Jiang, H. S., Gu, S. Z., et al., 2016. Cisuralian-Guadalupian Conodont Sequence from the Shaiwa Section, Ziyun, Guizhou, South China. Palaeogeography, Palaeoclimatology, Palaeoecology, 457: 1-22. https://doi.org/10.1016/j.palaeo.2016.05.030 |
| Wang, Z. H., 1978. Permian-Lower Triassic Conodonts of the Liangshan Area, Southern Shaanxi. Acta Palaeontologica Sinica, 17(2): 213-227 (in Chinese with English Abstract) http://www.researchgate.net/publication/312840646_Permian-Lower_Triassic_conodonts_of_the_Liangshan_area_southern_Shaanxi |
| Wu, Q., Ramezani, J., Zhang, H., et al., 2017. Calibrating the Guadalupian Series (Middle Permian) of South China. Palaeogeography, Palaeoclimatology, Palaeoecology, 466: 361-372. https://doi.org/10.1016/j.palaeo.2016.11.011 |
| Yao, Y., Yan, J. X., Li, A. Z., 2012. Sedimentary Features and Evolution of Mid-Permian Carbonates from Laibin of Guangxi. Earth Science-Journal of China University of Geosciences, 37(2): 184-194 (in Chinese with English Abstract) |
| Yin, H. F., Huang, D. H., 1995. The Early Palaoeozoic Zhan'an-Xichuan Block and the Evolution of the Small Qinling Archipelagic Ocean Basin. Acta Geologica Sinica, 69(3): 193-204 (in Chinese with English Abstract) http://epub.cnki.net/grid2008/docdown/docdownload.aspx?filename=DZXE199503000&dbcode=CJFD&year=1995&dflag=pdfdown |
| Yin, H. F., Peng, Y. Q., 1995. Evolution of the Phanerozoc Paleo-Ocean of Qinling. Earth Science-China University of Geosciences, 20(6): 605-611 (in Chinese with English Abstract) http://www.researchgate.net/publication/313618942_Evolution_of_the_Phanerozoic_paleo-ocean_of_Qinling |
| Yin, H. F., Zhang, K. X., 1998. Evolution and Characteristics of the Central Orogenic Belt. Earth Science-China University of Geosciences, 23(5): 438-442 (in Chinese with English Abstract) http://ci.nii.ac.jp/naid/10026542178 |
| Youngquist, W. L., Miller, A. K., 1949. Conodonts from the Late Mississippian Pella Beds of South-Central Iowa. Journal of Paleontology, 23(6): 617-622. https://doi.org/10.2307/1299669 |
| Yuan, D. X., Chen, J., Zhang, Y. C., et al., 2015. Changhsingian Conodont Succession and the End-Permian Mass Extinction Event at the Daijiagou Section in Chongqing, Southwest China. Journal of Asian Earth Sciences, 105(14): 234-251. https://doi.org/10.1016/j.jseaes.2015.04.002 |
| Zhang, G. W., Dong, Y. P., Lai, S. C., et al., 2004. Mianlüe Tectonic Zone and Mianlüe Suture Zone on Southern Margin of Qinling-Dabie Orogenic Belt. Science in China Series D, 47(4): 300-316. https://doi.org/10.1360/02yd0526 |
| Zhang, L. L., Zhang, N., Xia, W. C., 2007. Conodont Succession in the Guadalupian-Lopingian Boundary Interval (Upper Permian) of the Maoershan Section, Hubei Province, China. Micropaleontology, 53(6): 433-446. https://doi.org/10.2113/gsmicropal.53.6.433 |
| Zhang, N., Henderson, C. M., Xia, W. C., et al., 2010. Conodonts and Radiolarians through the Cisuralian-Guadalupian Boundary from the Pingxiang and Dachongling Sections, Guangxi Region, South China. Alcheringa, 34(2): 135-160. https://doi.org/10.1080/03115510903523292 |
| Zhao, Z., 1983. Conodont Sequences in Weixian and Xixia Stages of China. Xinjiang Petroleum Geology, 1: 38-61 (in Chinese) |
Figures(10) / Tables(2)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: