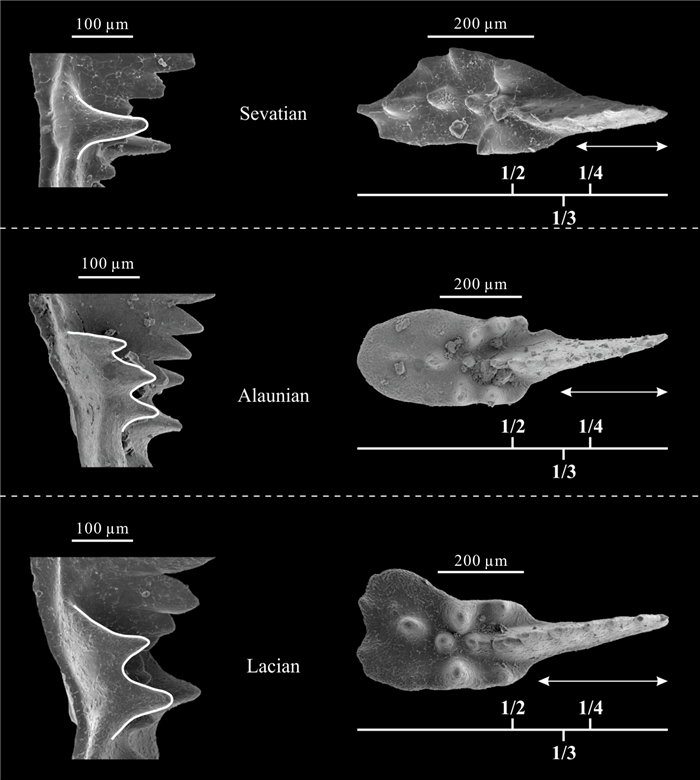
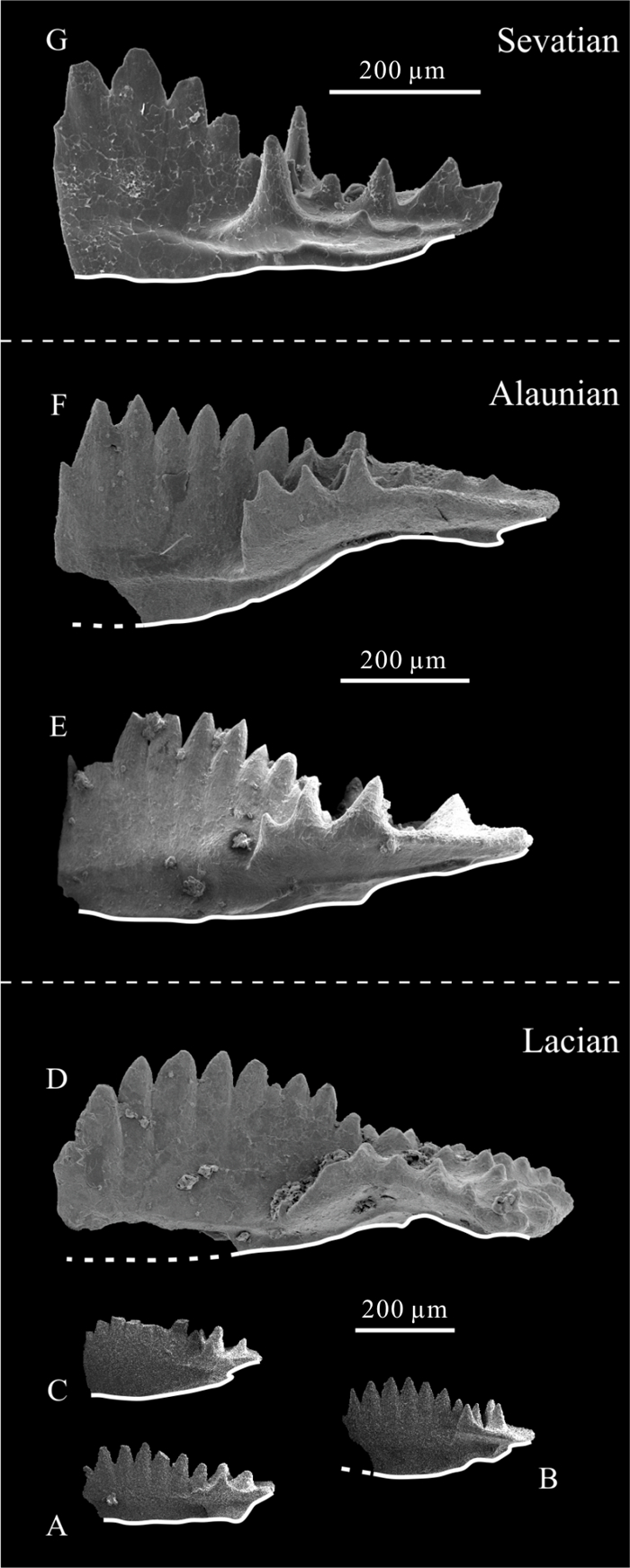
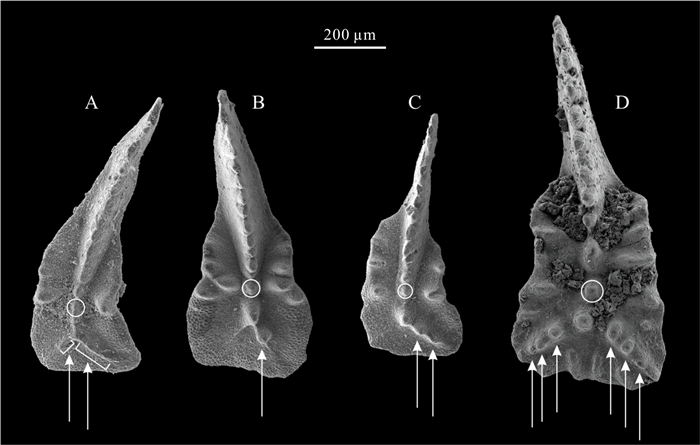
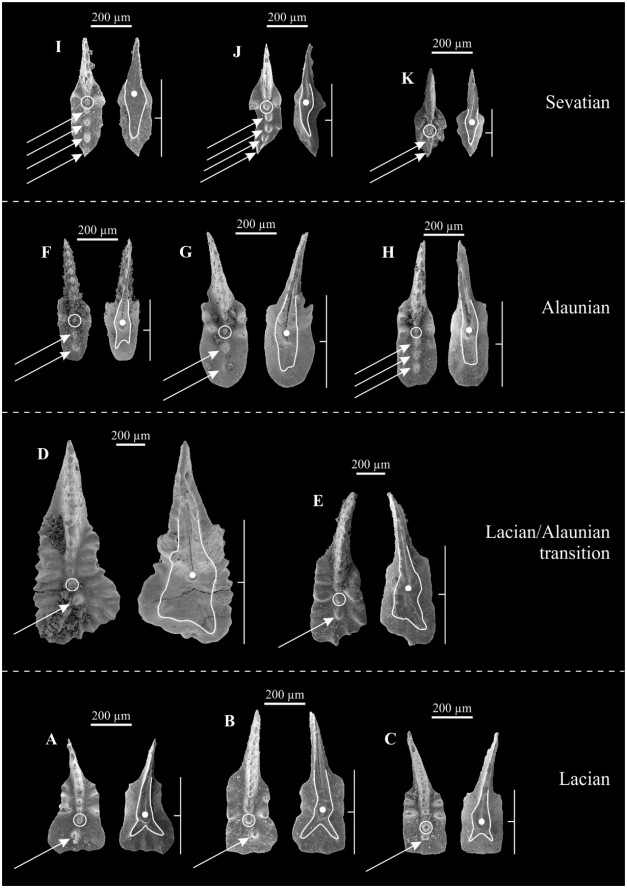
| Citation: | Viktor Karádi. Evolutionary Trends of the Genus Ancyrogondolella (Conodonta) and Related Taxa in the Norian (Late Triassic). Journal of Earth Science, 2021, 32(3): 700-708. doi: 10.1007/s12583-020-1381-z

|

| Baumgartner, N., Gawlick, H. -J., Missoni, S., et al., 2018. Open Marine Carnian Succession from the Apuseni Mountains (Codru-Moma Nappe System, Romania). In: Neubauer, F., Brendel, U., Friedl, G., eds., Advances of Geology in Southeast European Mountain Belts. XXI International Congress of the Carpathian Balkan Geological Association (CBGA). Sept. 10-13, 2018, Salzburg, Austria. Geologica Balcanica. 69 |
| Budurov, K., 1972. Ancyrogondolella triangularis gen. et sp. n. (Conodonta). Mitteilungen der Gesellschaft der Geologie-und Bergbaustudenten, 21: 853-860 |
| Budurov, K. J., Sudar, M. N., 1990. Late Triassic Conodont Stratigraphy. In: Ziegler, W., ed., 1st International Senckenberg Conference and 5th European Conodont Symposium (ECOS V) Contributions VI—Papers on Conodonts and Ordovician to Triassic Conodont Stratigraphy. Courier Forschungsinstitut Senckenberg, 118: 203-240 |
| Channell, J. E. T., Kozur, H. W., Sievers, T., et al., 2003. Carnian-Norian Biomagnetostratigraphy at Silická Brezová (Slovakia): Correlation to Other Tethyan Sections and to the Newark Basin. Palaeogeography, Palaeoclimatology, Palaeoecology, 191(2): 65-109. https://doi.org/10.1016/s0031-0182(02)006545 |
| Diakow, L., Orchard, M. J., Friedman, R., 2011. Absolute Ages for the Norian Stage: A Contribution from Southern British Columbia, Canada. In: Haggart, J. W., Smith, P. L., eds., Proceedings of the 21st Canadian Paleontology Conference. Canadian Paleontology Conference Proceedings, 9: 27-28 |
| Diakow, L., Orchard, M. J., Friedman, R., 2012. Absolute Ages for the Norian Stage: A Further Contribution from Southern British Columbia, Canada. Cordilleran Tectonics Workshop. Geological Association of Canada, Pacific Section, 2 |
| Du, Y. X., Bertinelli, A., Jin, X., et al., 2020. Integrated Conodont and Radiolarian Biostratigraphy of the Upper Norian in Baoshan Block, Southwestern China. Lethaia, 53: 533-545. https://doi.org/10.1111/let.12374 |
| Furin, S., Preto, N., Rigo, M., et al., 2006. High-Precision U-Pb Zircon Age from the Triassic of Italy: Implications for the Triassic Time Scale and the Carnian Origin of Calcareous Nannoplankton and Dinosaurs. Geology, 34: 1009-1012. https://doi.org/10.1130/g22967a.1 |
| Galbrun, B., Boulila, S., Krystyn, L., et al., 2020. "Short" or "Long" Rhaetian? Astronomical Calibration of Austrian Key Sections. Global and Planetary Change, 192: 103253. https://doi.org/10.1016/j.gloplacha.2020.103253 |
| Gehrels, G. E., Saleeby, J. B., Berg, H. C., 1987. Geology of Annette, Gravina, and Duke Islands, Southeastern Alaska. Canadian Journal of Earth Sciences, 24: 866-881 doi: 10.1139/e87-086 |
| Guenser, P., Souquet, L., Dolédec, S., et al., 2019. Deciphering the Roles of Environment and Development in the Evolution of a Late Triassic Assemblage of Conodont Elements. Paleobiology, 45(3): 440-457. https://doi.org/10.1017/pab.2019.14 |
| Huckriede, R., 1958. Die Conodonten der Mediterranen Trias und ihr Stratigraphischer Wert. Paläontologische Zeitschrift, 32(3): 141-175 doi: 10.1007/BF02989028.pdf |
| Karádi, V., 2018. Middle Norian Conodonts from the Buda Hills, Hungary: An Exceptional Record from the Western Tethys. Journal of Iberian Geology, 44: 155-174. https://doi.org/10.1007/s41513-017-0009-3 |
| Karádi, V., Kozur, H. W., Görög, Á., 2013. Stratigraphically Important Lower Norian Conodonts from the Csővár Borehole (Csv-1), Hungary— Comparison with the Conodont Succession of the Norian GSSP Candidate Pizzo Mondello (Sicily, Italy). In: Tanner, L. H., Spielmann, J. A., Lucas, S. G., eds., The Triassic System. New Mexico Museum of Natural History and Science Bulletin, 61: 284-295 |
| Karádi, V., Pelikán, P., Haas, J., 2016. Conodont Biostratigraphy of Upper Triassic Dolomites of the Buda Hills (Transdanubian Range, Hungary). Bulletin of the Hungarian Geological Society, 146(4): 371-386 (in Hungarian with English Abstract) http://www.researchgate.net/publication/316665083_Conodont_biostratigraphy_of_Upper_Triassic_dolomites_of_the_Buda_Hills_Transdanubian_Range_Hungary |
| Karádi, V., Kolar-Jurkovšek, T., Jurkovšek, B., 2019. The Lower/Middle Norian (Upper Triassic) Transition: Conodonts of the Dovško Succession, Slovenia. 3rd International Congress on Stratigraphy, July 2-5, 2019, Milano, Italy. Abstract Book. Società Geologica Italiana. 53. https://doi.org/10.3301/absgi.2019.04 |
| Karádi, V., Virág, A., Kolar-Jurkovšek, T., et al., 2020. Stress-Related Evolution in Triassic Conodonts and the Middle Norian Juvenile Mortality. In: Guex, J., Torday, J. S., Miller, W. B. Jr., eds., Morphogenesis, Environmental Stress and Reverse Evolution. Springer, Cham. 37-58. https://doi.org/10.1007/978-3-030-47279-5_4 |
| Karádi, V., Kolar-Jurkovšek, T., Gale, L., et al., 2021. New Advances in Biostratigraphy of the Lower/Middle Norian Transition: Conodonts of the Dovško Section, Slovenia. Journal of Earth Science, 32(3): 677-699. https://doi.org/10.1007/s12583-020-1382-y |
| Kent, D. V., Olsen, P. E., Muttoni, G., 2017. Astrochronostratigraphic Polarity Time Scale (APTS) for the Late Triassic and Early Jurassic from Continental Sediments and Correlation with Standard Marine Stages. Earth-Science Reviews, 166: 153-180. https://doi.org/10.1016/j.earscirev.2016.12.014 |
| Krystyn, L., 2003. Upper Triassic Substage Boundaries and Their Ammonoid Record: Divided between Gradation, Faunal Turnover and Extinction. Joint Annual Meeting GAC-MAC-SEG, Vancouver |
| Krystyn, L., Richoz, S., Gallet, Y., et al., 2007. Updated Bio-and Magnetostratigraphy from Steinbergkogel (Austria), Candidate GSSP for the Base of the Rhaetian Stage. Albertiana, 36: 164-173 http://www.researchgate.net/publication/46697632_Updated_bio-_and_magnetostratigraphy_from_Steinbergkogel_Austria_candidate_GSSP_for_the_base_of_the_Rhaetian_stage |
| Maron, M., Rigo, M., Bertinelli, A., et al., 2015. Magnetostratigraphy, Biostratigraphy, and Chemostratigraphy of the Pignola-Abriola Section: New Constraints for the Norian-Rhaetian Boundary. Geological Society of America Bulletin, 127(7/8): 962-974. https://doi.org/10.1130/b31106.1 |
| Mazza, M., Rigo, M., Nicora, A., 2011. A New Metapolygnathus Platform Conodont Species and Its Implications for Upper Carnian Global Correlation. Acta Palaeontologica Polonica, 56(1): 121-131. https://doi.org/10.4202/app.2009.1104 |
| Mazza, M., Rigo, M., Gullo, M., 2012a. Taxonomy and Biostratigraphic Record of the Upper Triassic Conodonts of the Pizzo Mondello Section (Western Sicily, Italy), GSSP Candidate for the Base of the Norian. Rivista Italiana di Paleontologia e Stratigrafia, 118(1): 85-130. https://doi.org/10.13130/2039-4942/5993 |
| Mazza, M., Cau, A., Rigo, M., 2012b. Application of Numerical Cladistics Analyses to the Carnian-Norian Conodonts: New Approach for Phylogenetic Interpretations. Journal of Systematic Palaeontology, 10(3): 401-422. https://doi.org/10.1080/14772019.2011.573584 |
| Mazza, M., Nicora, A., Rigo, M., 2018. Metapolygnathus parvus Kozur, 1972 (Conodonta): A Potential Primary Marker for the Norian GSSP (Upper Triassic). Bollettino della Società Paleontologica Italiana, 57(2): 81-101 http://www.researchgate.net/publication/327836318_Metapolygnathus_parvus_Kozur_1972_Conodonta_A_potential_primary_marker_for_the_Norian_GSSP_Upper_Triassic |
| Moix, P., Kozur, H. W., Stampfli, G. M., et al., 2007. New Paleontological, Biostratigraphic and Paleogeographic Results from the Triassic of the Mersin Mélange, SE Turkey. New Mexico Museum of Natural History and Science Bulletin, 41: 282-311 http://www.researchgate.net/publication/286284961_New_paleontological_biostratigraphic_and_paleogeographic_results_from_the_Triassic_of_the_Mersin_Melange_SE_Turkey |
| Mosher, L. C., 1968. Triassic Conodonts from Western North America and Europe and Their Correlation. Journal of Paleontology, 42(4): 895-946 |
| Noyan, Ö., Kozur, H., 2007. Revision of the Late Carnian-Early Norian Conodonts from the Stefanion Section (Argolis, Greece) and Their Paleobiogeographic Implications. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 245(2): 159-178. https://doi.org/10.1127/0077-7749/2007/0245-0159 |
| Noyan, Ö. F., Vrielynck, B., 2000. Importance of Morphogenetic Analysis in Taxonomy: An Example from Triassic Platform Conodonts. Neues Jahrbuch für Geologie und Paläontologie-Monatshefte, 10(11): 577-594 http://www.researchgate.net/publication/296720379_Importance_of_morphogenetic_analysis_in_taxonomy_an_example_from_Triassic_platform_conodonts |
| Ogg, J. G., 2012. Triassic. In: Gradstein, F. M., Ogg, J. G., Schmitz, M. D., et al., eds., The Geological Time Scale. Elsevier, Amsterdam. 681-730 |
| Orchard, M. J., 1983. Epigondolella Populations and Their Phylogeny and Zonation in the Upper Triassic. Fossils and Strata, 15: 177-192 http://www.researchgate.net/publication/285799635_Epigondolella_populations_and_their_phylogeny_and_zonation_in_the_Upper_Triassic |
| Orchard, M. J., 1991a. Late Triassic Conodont Biochronology and Biostratigraphy of the Kunga Group, Queen Charlotte Islands, British Columbia. Geological Survey of Canada Paper, 90-10: 173-193 http://ci.nii.ac.jp/naid/10021006710 |
| Orchard, M. J., 1991b. Upper Triassic Conodont Biochronology and New Index Species from the Canadian Cordillera. In: Orchard, M. J., McCracken, A. D., eds., Ordovician to Triassic Conodont Paleontology of the Canadian Cordillera. Geological Survey of Canada Bulletin, 417: 299-335 |
| Orchard, M. J., 1994. Late Triassic (Norian) Conodonts from Peru. Palaeontographica Abteilung A, 233(1-6): 203-208 http://www.schweizerbart.de/papers/pala/detail/A233/71552/Late_Triassic_Norian_conodonts_from_Peru |
| Orchard, M. J., 2006. Late Paleozoic and Triassic Conodont Faunas of Yukon and Northern British Columbia and Implications for the Evolution of the Yukon-Tanana Terrane. In: Colpron, M., Nelson, J. L., eds., Paleozoic Evolution and Metallogeny of Pericratonic Terranes at the Ancient Pacific Margin of North America, Canadian and Alaskan Cordillera. Geological Association of Canada Special Paper, 45: 229-260 |
| Orchard, M. J., 2014. Conodonts from the Carnian-Norian Boundary (Upper Triassic) of Black Bear Ridge. New Mexico Museum of Natural History and Science Bulletin, 64: 1-139 http://www.researchgate.net/publication/292234849_Conodonts_from_the_Carnian-Norian_boundary_Upper_Triassic_of_Black_Bear_Ridge_Northeasern_British_Columbia_Canada |
| Orchard, M. J., 2018. The Lower-Middle Norian (Upper Triassic) Boundary: New Conodont Taxa and a Refined Biozonation. In: Over, D. J., Henderson, C. M., eds., Conodont Studies Dedicated to the Careers and Contributions of Anita Harris, Glen Merrill, Carl Rexroad, Walter Sweet, and Bruce Wardlaw. Bulletins of American Paleontology, 395/396: 165-193 |
| Rigo, M., Mazza, M., Karádi, V., et al., 2018. New Upper Triassic Conodont Biozonation of the Tethyan Realm. In: Tanner, L. H., ed., The Late Triassic World: Earth in a Time of Transition. Topics in Geobiology, 46: 189-235. https://doi.org/10.1007/978-3-319-68009-5_6 |
| Roniewicz, E., 2011. Early Norian (Triassic) Corals from the Northern Calcareous Alps, Austria, and the Intra-Norian Faunal Turnover. Acta Palaeontologica Polonica, 56(2): 401-428. https://doi.org/10.4202/app.2009.0092 |
| Wotzlaw, J.-F., Guex, J., Bartolini, A., et al., 2014. Towards Accurate Numerical Calibration of the Late Triassic: High-Precision U-Pb Geochronology Constraints on the Duration of the Rhaetian. Geology, 42(7): 571-574. https://doi.org/10.1130/g35612.1 |
Figures(5)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: