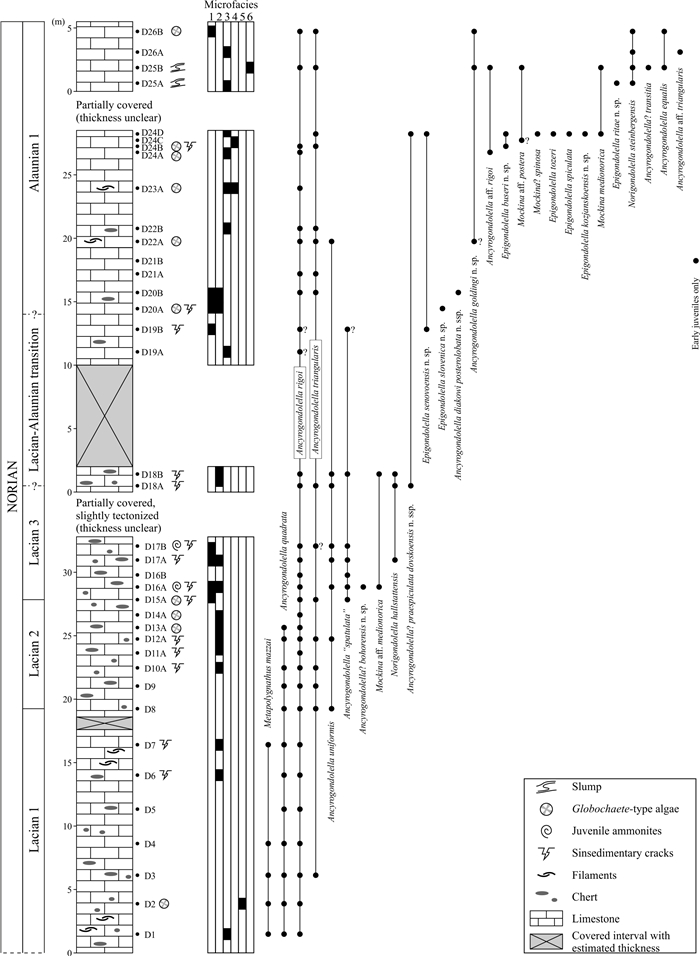
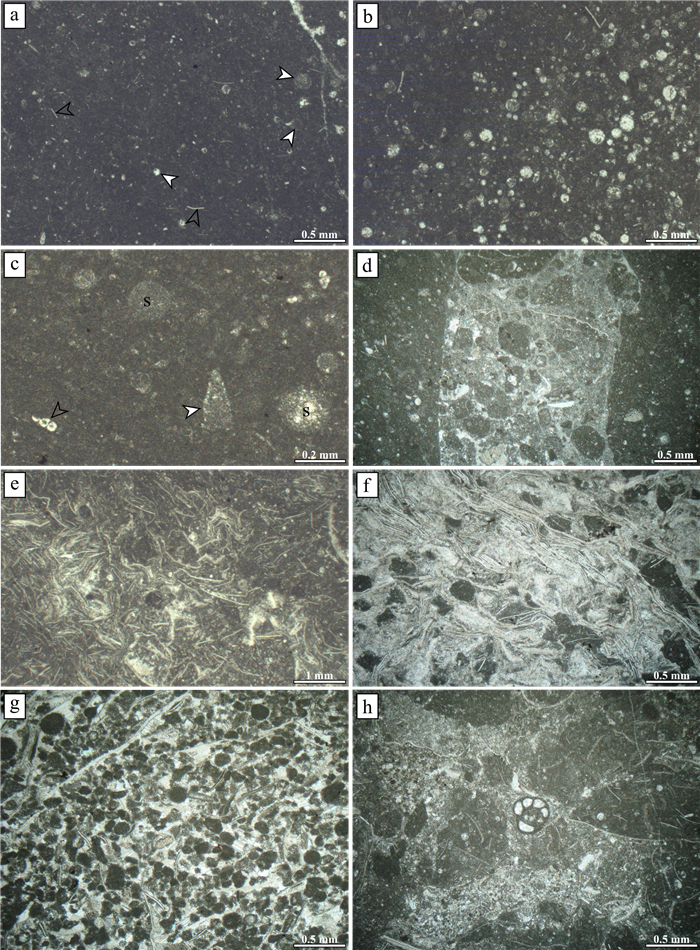
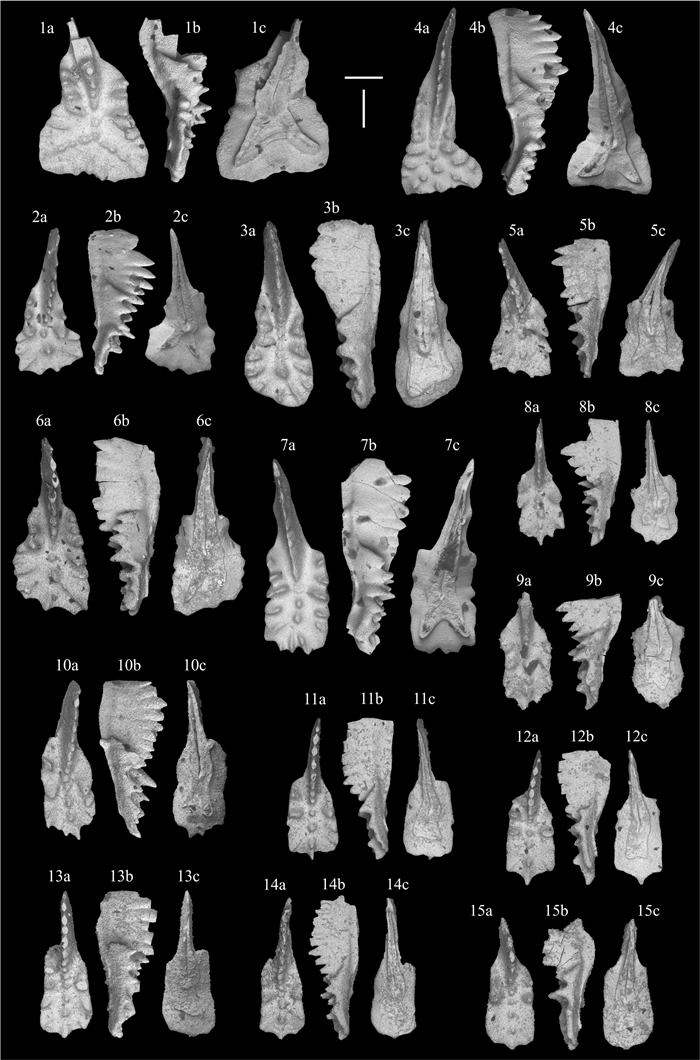
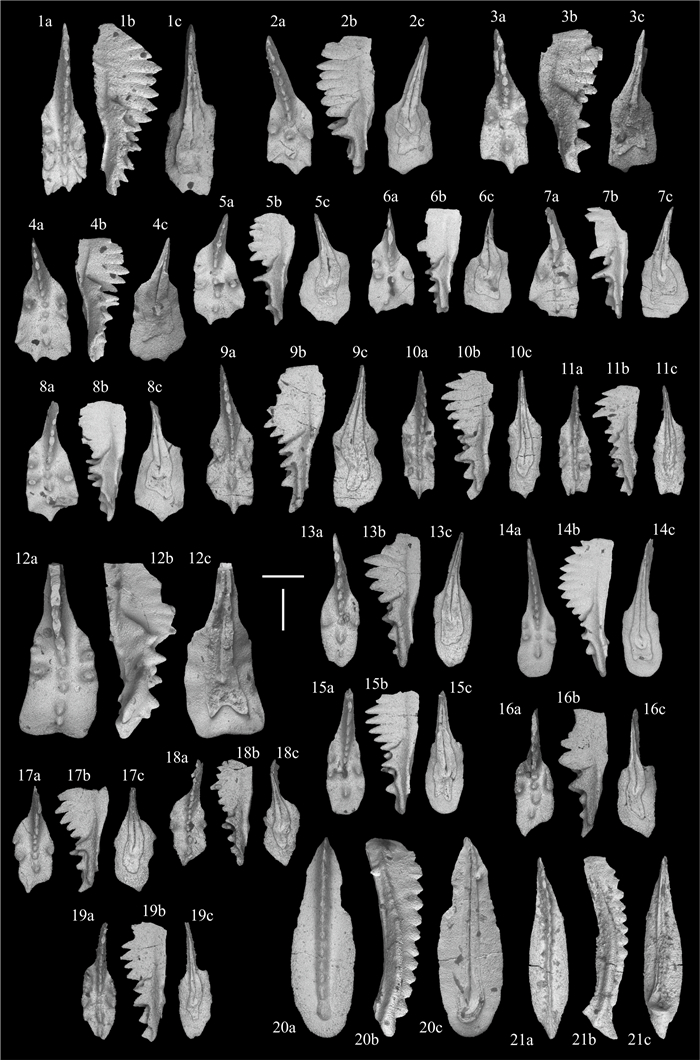
| Citation: | Viktor Karádi, Tea Kolar-Jurkovšek, Luka Gale, Bogdan Jurkovšek. New Advances in Biostratigraphy of the Lower/Middle Norian Transition: Conodonts of the Dovško Section, Slovenia. Journal of Earth Science, 2021, 32(3): 677-699. doi: 10.1007/s12583-020-1382-y

|

| Balini, M., Bertinelli, A., Di Stefano, P., et al., 2010. The Late Carnian-Rhaetian Succession at Pizzo Mondello (Sicani Mountains). Albertiana, 39: 36-57 http://www.researchgate.net/publication/260640296_The_Late_Carnian-Rhaetian_succession_at_Pizzo_Mondello_Sicani_Mountains |
| Budurov, K., 1972. Ancyrogondolella triangularis gen. et sp. n. (Conodonta). Mitteilungen der Gesellschaft der Geologie-und Bergbaustudenten, 21: 853-860 |
| Buser, S., 1978. Basic Geological Map SFRY, Sheet 33-67, Celje. Federal Geological Survey, Beograd |
| Buser, S., 1986. Explanatory Book to Basic Geological Map SFRY, Sheet Tolmin and Videm (Udine). Federal Geological Survey, Beograd. 103 |
| Buser, S., 1989. Development of the Dinaric and the Julian Carbonate Platforms and of the Intermediate Slovenian Basin (NW Yugoslavia). Bollettino della Società Geologica Italiana, 40: 313-320 http://www.researchgate.net/publication/285475171_Development_of_the_Dinaric_and_the_Julian_carbonate_platforms_and_of_the_intermediate_Slovenian_Basin_NW_Yugoslavia |
| Buser, S., 2009. Geological Map of Slovenia, Scale 1: 250 000. Geological Survey of Slovenia, Ljubljana |
| Buser, S., Kolar-Jurkovšek, T., Jurkovšek, B., 2008. The Slovenian Basin during the Triassic in the Light of Conodont Data. Bollettino della Società Geologica Italiana, 127: 257-263 http://italianjgeo.geoscienceworld.org/content/127/3/257.abstract |
| Celarc, B., Kolar-Jurkovšek, T., 2008. The Carnian-Norian Basin-Platform System of the Martuljek Mountain Group (Julian Alps, Slovenia): Progradation of the Dachstein Carbonate Platform. Geologica Carpathica, 59(3): 211-224 http://www.researchgate.net/publication/282821394_The_Carnian-Norian_basin-platform_system_of_the_Martuljek_Mountain_Group_Julian_Alps_Slovenia_Progradation_of_the_Dachstein_carbonate_platform/download |
| Channell, J. E. T., Kozur, H. W., Sievers, T., et al., 2003. Carnian-Norian Biomagnetostratigraphy at Silická Brezová (Slovakia): Correlation to Other Tethyan Sections and to the Newark Basin. Palaeogeography, Palaeoclimatology, Palaeoecology, 191: 65-109. https://doi.org/10.1016/s0031-0182(02)006545 |
| Cousin, M., 1981. Les Rapports Alpes-Dinarides; Les Confins de l'Italie et de Yougoslavie. Société Géologique du Nord, 5: 1-521 http://www.researchgate.net/publication/285798955_Les_rapports_Alpes-Dinarides |
| Dunham, R. J., 1962. Classification of Carbonate Rocks according to Depositional Texture. In: Ham, W. E., ed., Classification of Carbonate Rocks, a Symposium. American Association of Petroleum Geology Memoirs, 1: 108-121. https://doi.org/10.1306/m1357 |
| Dzik, J., 1976. Remarks on the Evolution of Ordovician Conodonts. Acta Palaeontologica Polonica, 21: 395-455 http://ci.nii.ac.jp/naid/10019474740 |
| Embry, A. F., Klovan, J. E., 1971. A Late Devonian Reef Tract on Northeastern Banks Island, N.W.T. Bulletin of Canadian Petroleum Geology, 19: 730-781 |
| Flügel, E., 2004. Microfacies of Carbonate Rocks: Analysis, Interpretation and Application. Springer-Verlag, Berlin Heidelberg. 976 |
| Gale, L., 2010. Microfacies Analysis of the Upper Triassic (Norian) "Bača Dolomite": Early Evolution of the Western Slovenian Basin (Eastern Southern Alps, Western Slovenia). Geologica Carpathica, 61: 293-308 doi: 10.2478/v10096-010-0017-0 |
| Gale, L., Kolar-Jurkovšek, T., Šmuc, A., et al., 2012. Integrated Rhaetian Foraminiferal and Conodont Biostratigraphy from the Slovenian Basin, Eastern Southern Alps. Swiss Journal of Geosciences, 105: 435-462. https://doi.org/10.1007/s00015-012-0117-1 |
| Gale, L., Celarc, B., Caggiati, M., et al., 2015. Paleogeographic Significance of Upper Triassic Basinal Succession of the Tamar Valley, Northern Julian Alps (Slovenia). Goelogica Carpathica, 66(4): 269-283. https://doi.org/10.1515/geoca-2015-0025 |
| Gale, L., Kolar-Jurkovšek, T., Karničnik, B., et al., 2019. Triassic Deep-Water Sedimentation in the Bled Basin, Eastern Julian Alps, Slovenia. Geologija, 62: 153-173. https://doi.org/10.5474/geologija.2019.007 |
| Gawlick, H. -J., Böhm, F., 2000. Sequence and Isotope Stratigraphy of Late Triassic Distal Periplatform Limestones from the Northern Calcareous Alps (Kälberstein Quarry, Berchtesgaden Hallstatt Zone). International Journal of Earth Sciences, 89: 108-129. https://doi.org/10.1007/s005310050320 |
| Hayashi, S., 1968. The Permian Conodonts in Chert of the Adoyama Formation, Ashio Mountains, Central Japan. Earth Science, 22: 63-77 http://ci.nii.ac.jp/naid/110007089757 |
| Huckriede, R., 1958. Die Conodonten der Mediterranen Trias und ihr Stratigraphischer Wert. Paläontologische Zeitschrift, 32(3): 141-175 doi: 10.1007/BF02989028.pdf |
| Ishida, K., Hirsch, F., 2001. Taxonomy and Faunal Affinity of Late Carnian-Rhaetian Conodonts in the Southern Chichibu Belt, Shikoku, SW Japan. Rivista Italiana di Paleontologia e Stratigrafia, 107(2): 227-250 |
| Jarc, S., Jerina, S., Miler, M., et al., 2017. Mineralogical and Geochemical Characteristics of Mudstones in the Jersovec Chert Deposit. Geologija, 60: 223-234. https://doi.org/10.5474/geologija.2017.016 |
| Ji, Z. -S., Yao, J. -X., Yang, X. -D., et al., 2003. Conodont Zonations of Norian in Lhasa Area, Xizang (Tibet) and Their Global Correlation. Acta Palaeontologica Sinica, 42(3): 382-392 (in Chinese with English Abstract) http://europepmc.org/abstract/CBA/538788 |
| Karádi, V., 2018. Middle Norian Conodonts from the Buda Hills, Hungary: An Exceptional Record from the Western Tethys. Journal of Iberian Geology, 44: 155-174. https://doi.org/10.1007/s41513-017-0009-3 |
| Karádi, V., 2021. Evolutionary Trends of the Genus Ancyrogondolella (Conodonta) and Related Taxa in the Norian (Late Triassic). Journal of Earth Science, 32(3): 700-708. https://doi.org/10.1007/s12583-020-1381-z |
| Karádi, V., Kozur, H. W., Görög, Á., 2013. Stratigraphically Important Lower Norian Conodonts from the Csővár Borehole (Csv-1), Hungary— Comparison with the Conodont Succession of the Norian GSSP Candidate Pizzo Mondello (Sicily, Italy). In: Tanner, L. H., Spielmann, J. A., Lucas, S. G., eds., The Triassic System. New Mexico Museum of Natural History and Science Bulletin, 61: 284-295 |
| Karádi, V., Pelikán, P., Haas, J., 2016. Conodont Biostratigraphy of Upper Triassic Dolomites of the Buda Hills (Transdanubian Range, Hungary). Bulletin of the Hungarian Geological Society, 146(4): 371-386 (in Hungarian with English Abstract) http://www.researchgate.net/publication/316665083_Conodont_biostratigraphy_of_Upper_Triassic_dolomites_of_the_Buda_Hills_Transdanubian_Range_Hungary |
| Karádi, V., Kolar-Jurkovšek, T., Jurkovšek, B., 2019. The Lower/Middle Norian (Upper Triassic) Transition: Conodonts of the Dovško Succession, Slovenia. 3rd International Congress on Stratigraphy, July 2-5, 2019, Milano, Italy. Abstract Book. Società Geologica Italiana. 53. https://doi.org/10.3301/absgi.2019.04 |
| Karádi, V., Virág, A., Kolar-Jurkovšek, T., et al., 2020. Stress-Related Evolution in Triassic Conodonts and the Middle Norian Juvenile Mortality. In: Guex, J., Torday, J. S., Miller, W. B. Jr., eds., Morphogenesis, Environmental Stress and Reverse Evolution. Springer, Cham, 37-58. https://doi.org/10.1007/978-3-030-47279-5_4 |
| Kolar-Jurkovšek, T., 1982. Conodonts from Amphiclina Beds and Bača Dolomite. Geologija, 25: 167-188 |
| Kolar-Jurkovšek, T., Jurkovšek, B., 2019. Conodonts of Slovenia. Geological Survey of Slovenia, Ljubljana. 259 |
| Kovács, S., Kozur, H., 1980. Stratigraphische Reichweite der Wichtigsten Conodonten (ohne Zahnreihenconodonten) der Mittel-und Obertrias. Geologisch-Paläontologische Mitteilungen Innsbruck, 10(2): 47-78 |
| Kozur, H., 1989a. Significance of Events in Conodont Evolution for the Permian and Triassic Stratigraphy. Courier Forschungsinstitut Senckenberg, 117: 385-408 http://www.researchgate.net/publication/284667350_Significance_of_events_in_conodont_evolution_for_the_Permian_and_Triassic_stratigraphy |
| Kozur, H., 1989b. The Taxonomy of the Gondolellid Conodonts in the Permian and Triassic. Courier Forschungsinstitut Senckenberg, 117: 409-469 http://ci.nii.ac.jp/naid/10030498714/ |
| Kozur, H., 1990. Norigondolella n. gen., eine neue Obertriassische Conodontengattung. Paläontologische Zeitschrift, 64(1): 125-132 doi: 10.1007/BF02985926 |
| Kozur, H., 2003. Integrated Ammonoid-, Conodont and Radiolarian Zonation of the Triassic. Hallesches Jahrbuch für Geowissenschaften, B25: 49-79 |
| Kozur, H., Mostler, H., 1971. Probleme der Conodontenforschung in der Trias. Geologisch-Paläontologische Mitteilungen Innsbruck, 1(4): 1-19 http://www.researchgate.net/publication/285105046_Probleme_der_Conodontenforschung_in_der_Trias |
| Krystyn, L., 1980. Triassic Conodont Localities of the Salzkammergut Region (Northern Calcareous Alps). Abhandlungen des Geologischen Bundesanstalt, Second European Conodont Symposium, Guidebook and Abstracts. 61-98 |
| Krystyn, L., 2003. Upper Triassic Substage Boundaries and Their Ammonoid Record: Divided between Gradation, Faunal Turnover and Extinction. Joint Annual Meeting GAC-MAC-SEG, Vancouver |
| Krystyn, L., 2008. Excursion 1: The Hallstatt Pelagics—Norian and Rhaetian Fossillagerstaetten of Hallstatt. In: Krystyn, L., Mandl, G. W., eds., Upper Triassic Subdivisions, Zonations and Events. Berichte der Geologischen Bundesanstalt, 76: 81-98 |
| Krystyn, L., Mandl, G. W., Schauer, M., 2009. Growth and Termination of the Upper Triassic Platform Margin of the Dachstein Area (Northern Calcareous Alps, Austria). Austrian Journal of Earth Sciences, 102: 23-33 http://search.ebscohost.com/login.aspx?direct=true&db=aph&AN=43900081&site=ehost-live |
| Lindström, M., 1970. A Suprageneric Taxonomy of the Conodonts. Lethaia, 3: 427-445 doi: 10.1111/j.1502-3931.1970.tb00834.x |
| Mao, L., Tian, C., 1987. Late Triassic Conodonts from the Uppermost Mailonggang Formation in Mailonggang Village of Lhünzhub County, Xizang (Tibet), China. Bulletin of the Chinese Academy of Geological Sciences, 17: 159-168 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-DQXB198703012.htm |
| Mazza, M., Rigo, M., Gullo, M., 2012. Taxonomy and Biostratigraphic Record of the Upper Triassic Conodonts of the Pizzo Mondello Section (Western Sicily, Italy), GSSP Candidate for the Base of the Norian. Rivista Italiana di Paleontologia e Stratigrafia, 118(1): 85-130. https://doi.org/10.13130/2039-4942/5993 |
| Moix, P., Kozur, H. W., Stampfli, G. M., et al., 2007. New Paleontological, Biostratigraphic and Paleogeographic Results from the Triassic of the Mersin Mélange, SE Turkey. In: Lucas, S. G., Spielmann J. A., eds., The Global Triassic. New Mexico Museum of Natural History and Science Bulletin, 41: 282-311 |
| Mosher, L. C., 1968. Triassic Conodonts from Western North America and Europe and Their Correlation. Journal of Paleontology, 42(4): 895-946 |
| Muttoni, G., Mazza, M., Mosher, D., et al., 2014. A Middle-Late Triassic (Ladinian-Rhaetian) Carbon and Oxygen Isotope Record from the Tethyan Ocean. Palaeogeography, Palaeoclimatology, Palaeoecology, 399: 246-259. https://doi.org/10.1016/j.palaeo.2014.01.018 |
| Noyan, Ö., Kozur, H., 2007. Revision of the Late Carnian-Early Norian Conodonts from the Stefanion Section (Argolis, Greece) and Their Paleobiogeographic Implications. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 245(2): 159-178. https://doi.org/10.1127/0077-7749/2007/0245-0159 |
| Ogorelec, B., Dozet, S., 1997. Upper Triassic, Jurassic and Lower Cretaceous Beds in Eastern Sava Folds—Section Laze at Boštanj (Slovenia). Mining and Metallurgy Quarterly, 44: 223-235 |
| Orchard, M. J., 1991a. Upper Triassic Conodont Biochronology and New Index Species from the Canadian Cordillera. In: Orchard, M. J., McCracken A. D., eds., Ordovician to Triassic Conodont Paleontology of the Canadian Cordillera. Geological Survey of Canada Bulletin, 417: 299-335 |
| Orchard, M. J., 1991b. Late Triassic Conodont Biochronology and Biostratigraphy of the Kunga Group, Queen Charlotte Islands, British Columbia. In: Woodsworth, G. W., ed., Evolution and Hydrocarbon Potential of the Queen Charlotte Basin, British Columbia. Geological Survey of Canada, Paper 90-10: 173-193 |
| Orchard, M. J., 2006. Late Paleozoic and Triassic Conodont Faunas of Yukon and Northern British Columbia and Implications for the Evolution of the Yukon-Tanana Terrane. In: Colpron, M., Nelson, J. L., eds., Paleozoic Evolution and Metallogeny of Pericratonic Terranes at the Ancient Pacific Margin of North America, Canadian and Alaskan Cordillera. Geological Association of Canada Special Paper, 45: 229-260 |
| Orchard, M. J., 2014. Conodonts from the Carnian-Norian Boundary (Upper Triassic) of Black Bear Ridge. New Mexico Museum of Natural History and Science Bulletin, 64: 1-139 http://www.researchgate.net/publication/292234849_Conodonts_from_the_Carnian-Norian_boundary_Upper_Triassic_of_Black_Bear_Ridge_Northeasern_British_Columbia_Canada |
| Orchard, M. J., 2018. The Lower-Middle Norian (Upper Triassic) Boundary: New Conodont Taxa and a Refined Biozonation. In: Over, D. J., Henderson, C. M., eds., Conodont Studies Dedicated to the Careers and Contributions of Anita Harris, Glen Merrill, Carl Rexroad, Walter Sweet, and Bruce Wardlaw. Bulletins of American Paleontology, 395/396: 165-193 |
| Orchard, M. J., Whalen, P. A., Carter, E. S., et al., 2007. Latest Triassic Conodonts and Radiolarian-Bearing Successions in Baja California Sur. In: Lucas, S. G., Spielmann, J. A., eds., The Global Triassic. New Mexico Museum of Natural History and Science Bulletin, 41: 355-365 |
| Pálfy, J., Demény, A., Haas, J., et al., 2007. Triassic-Jurassic Boundary Events Inferred from Integrated Stratigraphy of the Csővár Section, Hungary. Palaeogeography, Palaeoclimatology, Palaeoecology, 244: 11-33. https://doi.org/10.1016/j.palaeo.2006.06.021 |
| Placer, L., 1999a. Contribution to the Macrotectonic Subdivision of the Border Region between Southern Alps and External Dinarides. Geologija, 41: 223-255 http://www.oalib.com/paper/2151645 |
| Placer, L., 1999b. Structural Meaning of the Sava folds. Geologija, 41: 191-221 http://www.oalib.com/paper/2151680 |
| Poljak, M., 2007. Structural-Tectonic Map of Slovenia in Scale 1: 250 000. Geological Survey of Slovenia, Ljubljana. 52 |
| Rigo, M., Mazza, M., Karádi, V., et al., 2018. New Upper Triassic Conodont Biozonation of the Tethyan Realm. In: Tanner, L. H., ed., The Late Triassic World: Earth in a Time of Transition. Topics in Geobiology, 46: 189-235. https://doi.org/10.1007/978-3-319-68009-5_6 |
| Roniewicz, E., 2011. Early Norian (Triassic) Corals from the Northern Calcareous Alps, Austria, and the Intra-Norian Faunal Turnover. Acta Palaeontologica Polonica, 56(2): 401-428. https://doi.org/10.4202/app.2009.0092 |
| Rožič, B., 2016. Paleogeographic Units. In: Novak, M., Rman, N., eds., Geological Atlas of Slovenia. Geological Survey of Slovenia, Ljubljana. 14-15 |
| Rožič, B., Kolar-Jurkovšek, T., Šmuc, A., 2009. Late Triassic Sedimentary Evolution of Slovenian Basin (Eastern Southern Alps): Description and Correlation of the Slatnik Formation. Facies, 55: 137-155. https://doi.org/10.1007/s10347-008-0164-2 |
| Rožič, B., Gale, L., Kolar-Jurkovšek, T., 2013. Extent of the Upper Norian— Rhaetian Slatnik Formation in the Tolmin Nappe, eastern Southern Alps. Geologija, 56: 175-186. https://doi.org/10.5474/geologija.2013.011 |
| Spatzenegger, A., 2011. Ammonites from the Zone of Cyrtopleurites Bicrenatus, Nor/Alaunium1 of the Hallstatt limestone. The Fossil Forum. http://www.thefossilforum.com/index.php?/blogs/entry/115-ammonites-from-the-zone-of-cyrtopleurites-bicrenatus/ |
| Trotter, J. A., Williams, I. A., Nicora, A., et al., 2015. Long-Term Cycles of Triassic Climate Change: A New δ18O Record from Conodont Apatite. Earth and Planetary Science Letters, 415: 165-174. https://doi.org/10.1016/j.epsl.2015.01.038 |
| Vrabec, M., Fodor, L., 2006. Late Cenozoic Tectonics of Slovenia: Structural Styles at the Northeastern Corner of the Adriatic Microplate. In: Pinter, N., Grenerczy, G., Weber, J., et al., eds., The Adria Microplate: GPS Geodesy, Tectonics and Hazards. NATO Science Series, IV, Earth and Environmental Sciences, 61: 151-168 |
| Vrielynck, B., 1987. Conodontes du Trias Perimediterraneen. Systematique, Stratigraphie. Documents des Laboratoires de Géologie Lyon, 97: 1-301 http://www.museocivico.rovereto.tn.it/patrimonio_abbonamenti_intermedio.jsp?area=69&ID_LINK=111098&page=1566&id_context=77666 |
Figures(7) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: