| Citation: | Ali Murat Kılıç. Anisian (Middle Triassic) Conodonts of the Kocaeli Triassic, Western Turkey. Journal of Earth Science, 2021, 32(3): 616-632. doi: 10.1007/s12583-020-1384-9

|

| Agematsu, S., Orchard, M. J., Sashida, K., 2008. Reconstruction of an Apparatus of Neostrachanognathus tahoensis from Oritate, Japan and Species of Neostrachanognathus from Oman. Palaeontology, 51(5): 1201-1211. https://doi.org/10.1111/j.1475-4983.2008.00804.x |
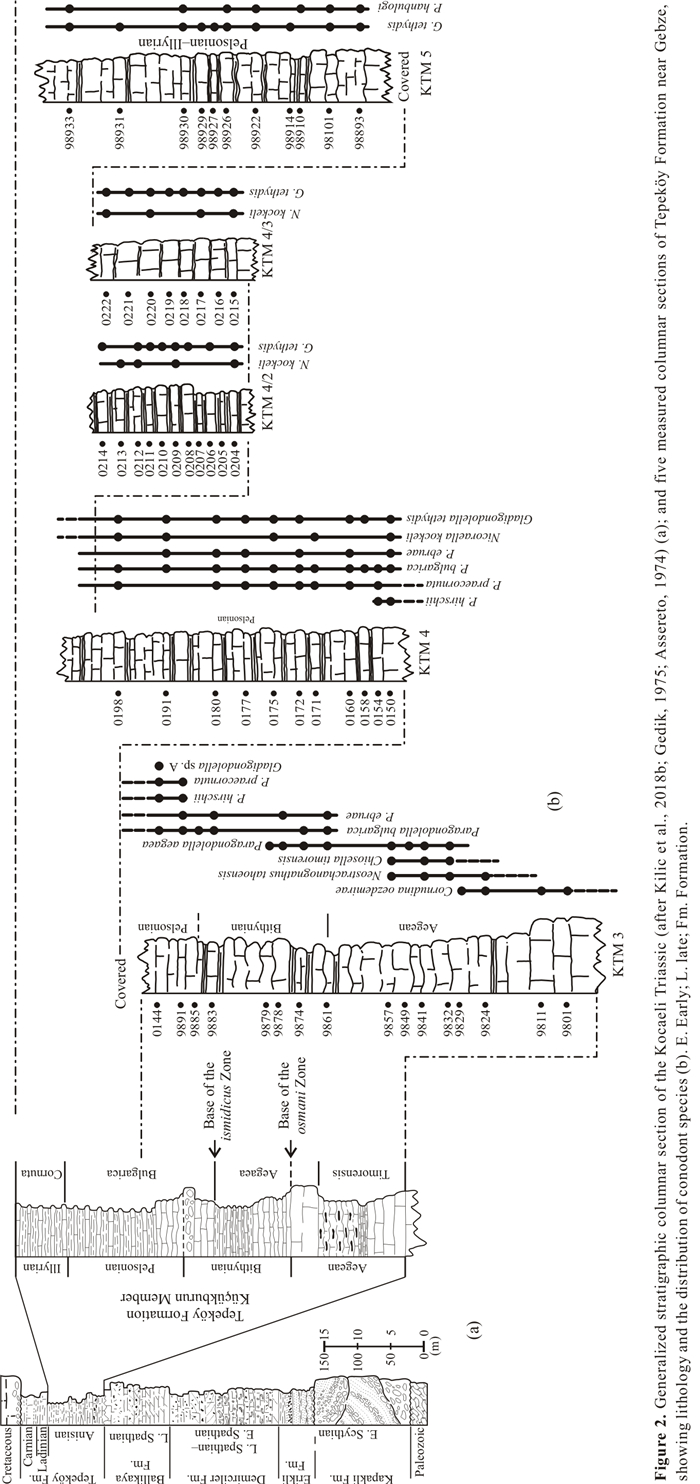
| Assereto, R., 1972. Notes on the Anisian Biostratigraphy of the Gebze Area (Kocaeli Peninsula, Turkey). Zeitschrift der Deutschen Geologischen Gesellschaft, 123(2): 435-444. https://doi.org/10.1127/zdgg/123/1972/435 |
| Assereto, R., 1974. Aegean and Bithynian: Proposal for Two New Anisian Substages. In: Zapfe, H., ed., The Stratigraphy of the Alpine-Mediterranean Triassic. Springer, Vienna. 23-39. https://doi.org/10.1007/978-3-7091-4497-8_2 |
| Bender, H., 1970. Zur Gliederung der Mediterranen Trias II. Die Conodontenchronologie der Mediterranen Trias. Annales Geologiques des Pays Helleniques, 19: 465-540 http://ci.nii.ac.jp/naid/10020877413 |
| Bender, H., Stoppel, D., 1965. Perm-Conodonten. Geologisches Jahrbuch, 82: 331-364 http://ci.nii.ac.jp/naid/10012506981 |
| Budurov, K., 1960. Uber die Anwesenheit von Conodonten im Anis bei Granitovo, Bezirk Vidin. Bulg. Geol. Soc. Rev., 21: 77-80 (in Bulgarian with English Summary) |
| Budurov, K., 1973. Carinella n. gen. und Revision der Gattung Gladigondolella (Conodonta). Comptes Rendus de l'Academie Bulgare des Sciences, 26: 799-802 |
| Budurov, K., 1975. Paragondolella foliata sp. n. [Conodonta] von der Trias des Ostbalkans. Rev. Bulg. Geol. Soc., 36(1): 79-81 |
| Budurov, K., 1980. Conodont Stratigraphy of the Balkanide Triassic. Rivista Italiana di Paleontologia e Stratigrafia, 85: 767-780 |
| Budurov, K. J., Stefanov, S., 1972. Platform-Condonten und Ihre Zonen in der Mittleren Trias Bulgariens. Mitt. Ges. Geol. Bergabaustud, 21: 829-852 http://www.researchgate.net/publication/281043692_Platform-Condonten_und_ihre_Zonen_in_der_Mittleren_Trias_Bulgariens |
| Budurov, K. J., Stefanov, S., 1975. Neue Daten über die Conodonten-Chronologie der Balkaniden Mittleren Trias. Comptes Rendus de lʼAcademia Bulgare des Sciences, 28(6): 791-794 http://www.researchgate.net/publication/281069937_Neue_Daten_uber_die_Conodonten-Chronologie_der_Balkaniden_mittleren_Trias |
| Budurov, K. J., Sudar, M. N., 1990. Late Triassic Conodont Stratigraphy. In: Ziegler, W., ed., 1st International Senckenberg Conference and 5th European Conodont Symposium (ECOS V) Contributions IV. Papers on Conodonts and Ordovician to Triassic Conodont Stratigraphy. Courier Forschungsinstitut Senckenberg, 118: 203-240 |
| Budurov, K. J., Sudar, M. N., Gupta, V. J., 1988. Kashmirella a New Early Triassic Conodont Genus. Bulletin of Indian Geologists Association, 21: 107-112 |
| Buryi, G. I., 1989. Triassic Conodonts and Stratigraphy of Sikhote-Alin. FEB Academy of Sciences of the USSR, Vladivostok. 136 |
| Chen, Y. L., Krystyn, L., Orchard, M. J., et al., 2016. A Review of the Evolution, Biostratigraphy, Provincialism and Diversity of Middle and Early Late Triassic Conodonts. Papers in Palaeontology, 2(2): 235-263. https://doi.org/10.1002/spp2.1038 |
| Chen, Y. L., Scholze, F., Richoz, S., et al., 2019a. Middle Triassic Conodont Assemblages from the Germanic Basin: Implications for Multi-Element Taxonomy and Biogeography. Journal of Systematic Palaeontology, 17(5): 359-377. https://doi.org/10.1080/14772019.2018.1424260 |
| Chen, Y. L., Richoz, S., Krystyn, L., et al., 2019b. Quantitative Stratigraphic Correlation of Tethyan Conodonts across the Smithian-Spathian (Early Triassic) Extinction Event. Earth-Science Reviews, 195: 37-51. https://doi.org/10.1016/j.earscirev.2019.03.004 |
| Clark, D. L., Sweet, W. C., Bergstrom, S. M., et al., 1981. Conodonta. Part W, Miscellanea, Supplement 2. In: Robison, R. A., ed., Treatise on İnvertebrate Paleontology. Geological Society of America, Boulder, Colo., University of Kansas, Lawrence |
| Dagys, A. S., 1987. Boreal Trias. Institute of Geology and Geophysics, Academy of Sciences of the USSR Siberian Branch. Transactions у, 689: 131 |
| Donoghue, P. C. J., Purnell, M. A., Aldridge, R. J., et al., 2008. The Interrelationships of 'Complex' Conodonts (Vertebrata). Journal of Systematic Palaeontology, 6(2): 119-153. https://doi.org/10.1017/s1477201907002234 |
| Dzik, J., 1976. Remarks on the Evolution of Ordovician Conodonts. Acta Palaeontologica Polonica, 21(4): 395-455 http://ci.nii.ac.jp/naid/10019474740 |
| Farabegoli, E., Levanti, D., Perri, M. C., et al., 1984. M. Bivera Formation: An Atypical Middle Triassic "Rosso Ammonitico" Facies from Southern Alps (Italy). Giornale di Geologia, 46: 33-46 http://www.researchgate.net/publication/256461287_M_Bivera_Formation_an_atypical_Middle_Triassic_Rosso_Ammonitico_facies_from_Southern_Alps_Italy |
| Franz, M., Kaiser, S. I., Fischer, J., et al., 2015. Eustatic and Climatic Control on the Upper Muschelkalk Sea (Late Anisian/Ladinian) in the Central European Basin. Global and Planetary Change, 135: 1-27. https://doi.org/10.1016/j.gloplacha.2015.09.014 |
| Gaetani, M., Jacobshagen, V., Nicora, A., et al., 1992. The Early-Middle Triassic Boundary at Chios. Rivista Italiana di Paleontologia e Stratigrafia, 98(2): 181-204 http://www.researchgate.net/publication/292233097_The_early-middle_Triassic_boundary_at_Chios_Greece |
| Gedik, İ., 1975. Die Conodonten der Trias auf der Kocaeli Halbinsel[Turkei]. Palaeontographica Abteilung, [A]: 99-160 |
| Götz, A. E., Gast, S., 2010. Basin Evolution of the Anisian Peri-Tethys: Implications from Conodont Assemblages of Lower Muschelkalk Key Sections (Central Europe). Zeitschrift Der Deutschen Gesellschaft Für Geowissenschaften, 161(1): 39-49. https://doi.org/10.1127/1860-1804/2010/0161-0039 |
| Golding, M. L., Orchard, M. J., Zonneveld, J. P., et al., 2014. An Exceptional Record of the Sedimentology and Biostratigraphy of the Montney and Doig Formations in British Columbia. Bulletin of Canadian Petroleum Geology, 62(3): 157-176 |
| Golding, M. L., Orchard, M. J., Zonneveld, J. P., et al., 2015. Determining the Age and Depositional Model of the Doig Phosphate Zone in Northeastern British Columbia Using Conodont Biostratigraphy. Bulletin of Canadian Petroleum Geology, 63(2): 143-170. https://doi.org/10.2113/gscpgbull.63.2.143 |
| Golding, M. L., Orchard, M. J., 2018. Magnigondolella, a New Conodont Genus from the Triassic of North America. Journal of Paleontology, 92(2): 207-220. https://doi.org/10.1017/jpa.2017.123 |
| Goudemand, N., Orchard, M. J., Bucher, H., et al., 2011. Is Chiosella timorensis a Good Index for the Olenekian-Anisian Boundary?. In: Håkansson, E., Trotter, J. A., eds., Programme and Abstracts: The XVII International Congress on the Carboniferous and Permian. July 2011, Perth. 61 |
| Goudemand, N., Orchard, M. J., Bucher, H., et al., 2012. The Elusive Origin of Chiosella timorensis (Conodont Triassic). Geobios, 45(2): 199-207. https://doi.org/10.1016/j.geobios.2011.06.001 |
| Grãdinaru, E., Kozur, H., Nicora, A., et al., 2006. The Chiosella timorensis Lineage and Correlation of the Ammonoids and Conodonts around the Base of the Anisian in the GSSP Candidate at Desli Caira (North Dobrogea, Romania). Albertiana, 34: 34-38 http://www.researchgate.net/publication/273693960_The_Chiosella_timorensis_lineage_and_correlation_of_the_ammonoids_and_conodonts_around_the_base_of_the_Anisian_in_the_GSSP_candidate_at_Desli_Caira_North_Dobrogea_Romania |
| Hayashi, S., 1968. The Permian Conodonts in Chert of the Adayama Formation, Ashio Mountains, Central Japan. Earth Science (Chikyu Kagaku), 22(2): 63-77 http://ci.nii.ac.jp/naid/110007089757 |
| Hirsch, F., 1969. La Succession des Faunes de Conodontes dans les Couches de Passage de lʼAnisien Supérieur au Ladinien Inférieur des Alpes Orientales et Méridionales. Arch. Sc. Genève, 22(1): 83-90 (in French) |
| Hirsch, F., 1975. Lower Triassic Conodonts from Israel. Bull. Geol. Surv. Israel, 66: 39-48 http://www.researchgate.net/publication/285785296_Lower_Triassic_conodonts_from_Israel |
| Hirsch, F., 1994. Triassic Conodonts as Ecological and Eustatic Sensors. In: Embry, A. F., Beauchamp, B., Glass, D. J., eds., Pangea: Global Environments and Resources. Memoir of the Canadian Society of Petroleum Geologists, 17: 949-959 |
| Hirschmann, C., 1959. Uber Conodonten aus dem Oberen Muschelkalk des Thüringer Beckens. Freiberger Forschungshefte, Paläontologie, 76: 35-86 |
| Huang, J. Y., Hu, S. X., Zhang, Q. Y., et al., 2019a. Gondolelloid Multielement Conodont Apparatus (Nicoraella) from the Middle Triassic of Yunnan Province, Southwestern China. Palaeogeography, Palaeoclimatology, Palaeoecology, 522: 98-110. https://doi.org/10.1016/j.palaeo.2018.07.015 |
| Huang, J. Y., Martínez-Pérez, C., Hu, S. X., et al., 2019b. Middle Triassic Conodont Apparatus Architecture Revealed by Synchrotron X-Ray Microtomography. Palaeoworld, 28(4): 429-440. https://doi.org/10.1016/j.palwor.2018.08.003 |
| Huckriede, R., 1958. Die Conodonten der Mediterranen Trias und ihr Stratigraphischer Wert. Paläontologische Zeitschrift, 32(3/4): 141-175 doi: 10.1007/BF02989028.pdf |
| Ishida, K., Hirsch, F., 2011. The Triassic Conodonts of the NW Malayan Kodiang Limestone Revisited: Taxonomy and Paleogeographic Significance. Gondwana Research, 19(1): 22-36. https://doi.org/10.1016/j.gr.2010.05.008 |
| Ishii, K., Nogami, Y., 1966. Discovery of Triassic Conodonts from the So-Called Palaeozoic Limestone in Kedah, Malaya. Osaka City University, Journal Geosciences, 9: 92-98 http://ci.nii.ac.jp/naid/110000003654/en |
| Karádi, V., Dulai, A., 2016. Sándor Kovácsʼs Conodont Collection. Annales Musei Historico-Naturalis Hungarici, 108: 221-229 |
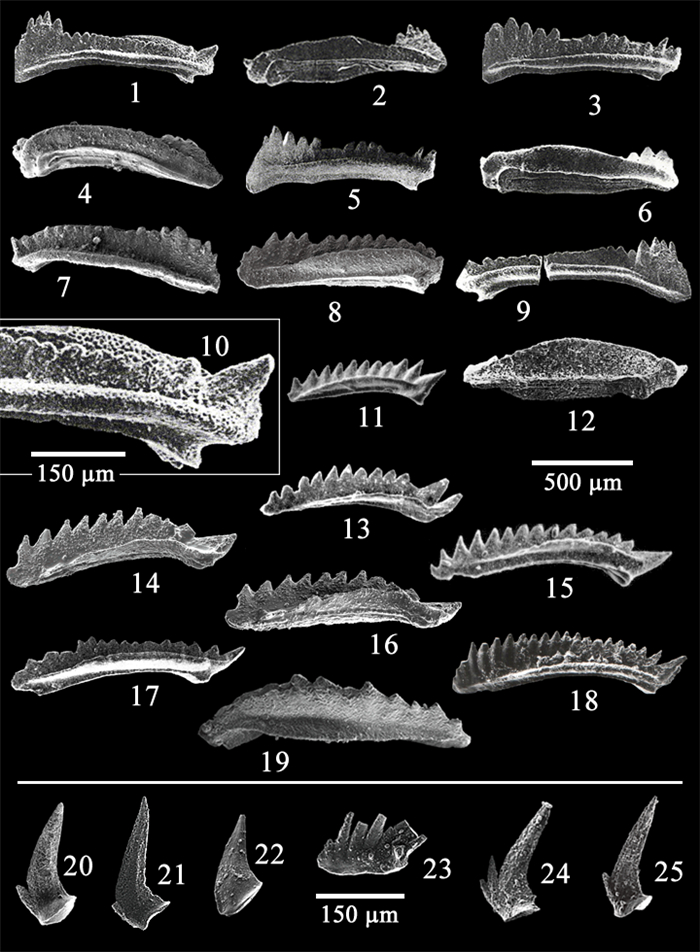
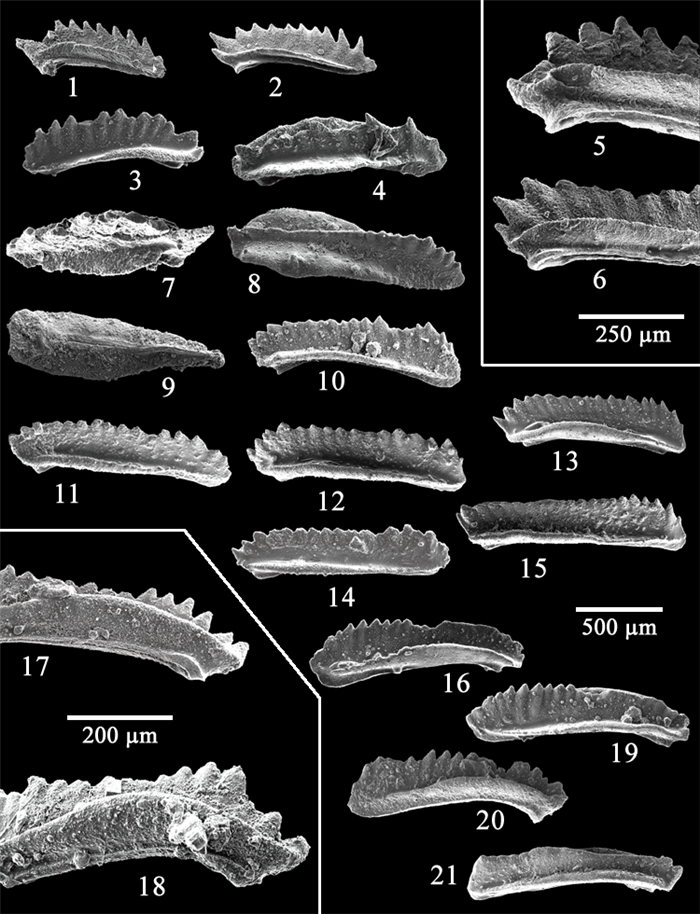
| Kılıç, A. M., 2004. Multielement Taxonomy of the Triassic Conodonts of the Kocaeli Region: [Dissertation]. Cumhuriyet University, Sivas, Turkey. 132 (in Turkish with English Abstract) |
| Kılıç, A. M., Plasencia, P., Guex, J., et al., 2018a. Challenging Darwin: Evolution of Triassic Conodonts and Their Struggle for Life in a Changing World. Stratigraphy and Timescales, 2: 333-389. https://doi.org/10.1016/bs.sats.2017.08.003 |
| Kılıç, A. M., Plasencia, P., Onder, F., 2018b. Debate on Skeletal Elements of the Triassic Conodont Cornudina Hirschmann. Acta Geologica Polonica, 68(2): 147-159 http://www.researchgate.net/publication/322759655_Debate_on_skeletal_elements_of_the_Triassic_conodont_Cornudina_Hirschmann |
| Klapper, G., Lindström, M., Sweet, W. C., et al., 1973. In: Ziegler, W., ed., Catalogue of Conodonts. Schweizerbart Science Publishers, Stuttgart. 1: XVIII |
| Koike, T., 1982. Review of Some Platform Conodonts of the Middle and Late Triassic of Japan. Science Reports of the Yokohama National University, Section 2: Biological and Geological, 2/29: 15-27 http://ci.nii.ac.jp/naid/110006151355/en |
| Koike, T., 1996. Skeletal Apparatuses of Triassic Conodonts of Cornudina. In: Noda, K., Sashida, K., eds., Professor Hisayoshi Igo Commemorative Volume on Geology and Paleontology of Japan and Southeast Asia. Gakujyutsu Tosho Insatsu Co. Ltd., Tokyo. 113-120 |
| Koike, T., 1998. Triassic Coniform Conodont Genera Aduncodina and Neostrachanognathus. Paleontological Research, 2(2): 120-129 http://ci.nii.ac.jp/naid/110002695175/en |
| Koike, T., 1999. Apparatus of a Triassic Conodont Species Cratognathodus multihamatus (Huckriede). Paleontological Research, 3(4): 234-248 http://ci.nii.ac.jp/naid/110002695215/en |
| Koike, T., 2016. Multielement Conodont Apparatuses of the Ellisoniidae from Japan. Paleontological Research, 20(3): 161-175. https://doi.org/10.2517/2016pr007 |
| Kolar-Jurkovšek, T., Jurkovšek, B., 2010. New Paleontological Evidence of the Carnian Strata in the Mežica Area (Karavanke Mts, Slovenia): Conodont Data for the Carnian Pluvial Event. Palaeogeography, Palaeoclimatology, Palaeoecology, 290(1/2/3/4): 81-88. https://doi.org/10.1016/j.palaeo.2009.06.015 |
| Kolar-Jurkovšek, T., Gaździcki, A., Jurkovšek, B., 2005. Conodonts and Foraminifera from the "Raibl Beds" (Carnian) of the Karavanke Mountains, Slovenia: Stratigraphical and Palaeobiological Implications. Geological Quarterly, 49(4): 429-438 |
| Kolar-Jurkovšek, T., Chen, Y. L., Jurkovšek, B., et al., 2017. Conodont Biostratigraphy of the Early Triassic in Eastern Slovenia. Paleontological Journal, 51(7): 687-703. https://doi.org/10.1134/s003103011707005x |
| Kovács, S., 2003. Pelsonian Conodonts from the Balaton Highland. Geologica Hungarica, Series Palaeontologica, 55: 159-177 http://www.researchgate.net/publication/238113910_The_Pelsonian_brachiopod_fauna_of_the_Balaton_Highland |
| Kozur, H. W., 1968. Neue Conodonten aus dem Oberen Muschelkalk des Germanischen Binnenbeckens. Monatsber. Deutsch. Akad. Wiss. Berlin, 10(2): 130-142, Pl. 1 |
| Kozur, H., 1980. Revision der Conodontenzonierung der Mittel-und Obertrias des Tethyalen Faunenreichs. Geologische-Paläontologische Mitteilungen Innsbruck, 10: 79-112 http://www.researchgate.net/publication/285105079_Revision_der_Conodontenzonierung_der_Mittel-_und_Obertrias_des_tethyalen_Faunenreichs |
| Kozur, H. W., 1989. The Taxonomy of the Gondolellids Conodonts in the Permian and Triassic. 1st International Senckenberg Conference and 5th European Conodont Symposium (ECOS V): Contributions III, Papers on Ordovician to Triassic Conodonts. 117: 409-469 |
| Kozur, H., 1993. Nicoraella postkockeli n. sp., a New Conodont Species from the Lower Carnian of Hungary. Neues Jahrbuch für Geologie und Paläontologie-Monatshefte, 7: 405-412. https://doi.org/10.1127/njgpm/1993/1993/405 |
| Kozur, H. W., 2003. Integrated Ammonoid-, Conodont and Radiolarian Zonation of the Triassic. Hall Jahrb. Abt. Geowiss. B, 25: 49-79 http://www.researchgate.net/publication/286181376_Integrated_ammonoid_conodont_and_radiolarian_zonation_of_the_Triassic_and_some_remarks_to_StageSubstage_subdivision_and_the_numeric_age_of_the_Triassic_stages |
| Kozur, H., Mock, R., 1991. New Middle Carnian and Rhaetian Conodonts from Hungary and the Alps. Stratigraphic Importance and Tectonic Implications for the Buda Mountains and Adjacent Areas. Jahrbuch der Geologischen Bundesanstalt, 134(2): 271-297 |
| Kozur, H. W., Mostler, H., 1970. Neue Conodonten aus der Trias. Ber. Nat. -Med. Verein Innsbruck, 58: 429-464 |
| Kozur, H. W., Mostler, H., 1971. Probleme der Conodontenforschung in der Trias. Geol. Palaont. Mitt. Innsbruck, 1(4): 1-19 http://www.researchgate.net/publication/285105046_Probleme_der_Conodontenforschung_in_der_Trias |
| Kozur, H. W., Mostler, H., 1972. Die Bedeutung der Conodonten fur Stratigraphische und Paleogeographische Untersuchungen in der Trias. Mitt. Ges. Geot. Bergbaustud., 21: 777-810 http://www.researchgate.net/publication/285105326_Die_Bedeutung_der_Conodonten_fur_stratigraphische_und_palaogeographische_Untersuchungen_in_der_Trias |
| Kozur, H. W., Mostler, H., 1992. Erster Paläontologischer Nachweis von Meliaticum und Sud-Rudabanyaicum in den Nordlichen Kalkalpen (Osterreich) und ihre Beziehung zu den Abfolgen den Westkarpaten. Geologische-Palaontologische Mitteilungen Innsbruck, 18: 87-129 |
| Lambert, L. L., Wardlaw, B. R., Henderson, C. M., 2007. Mesogondolella and Jinogondolella (Conodonta): Multielement Definition of the Taxa that Bracket the Basal Guadalupian (Middle Permian Series) GSSP. Palaeoworld, 16(1-3): 208-221. https://doi.org/10.1016/j.palwor.2007.05.017 |
| Lehrmann, D. J., Stepchinski, L., Altiner, D., et al., 2015. An Integrated Biostratigraphy (Conodonts and Foraminifers) and Chronostratigraphy (Paleomagnetic Reversals, Magnetic Susceptibility, Elemental Chemistry, Carbon Isotopes and Geochronology) for the Permian-Upper Triassic Strata of Guandao Section, Nanpanjiang Basin, South China. Journal of Asian Earth Sciences, 108: 117-135. https://doi.org/10.1016/j.jseaes.2015.04.030 |
| Lindström, M., 1970. A Suprageneric Taxonomy of the Conodonts. Lethaia, 3(4): 427-445. https://doi.org/10.1111/j.1502-3931.1970.tb00834.x |
| Márquez-Aliaga, A., Valenzuela-Ríos, J. I., Calvet, F., et al., 2000. Middle Triassic Conodonts from Northeastern Spain: Biostratigraphic Implications. Terra Nova, 12(2): 77-83. https://doi.org/10.1046/j.1365-3121.2000.00273.x |
| Mazza, M., Rigo, M., Gullo, M., 2012. Taxonomy and Biostratigraphic Record of the Upper Triassic Conodonts of the Pizzo Mondello Section (Western Sicily, Italy), GSSP Candidate for the Base of the Norian. Rivista Italiana di Paleontologia e Stratigrafia, 8: 85-130 |
| Meco, S., 1999. Conodont Biostratigraphy of Triassic Pelagic Strata, Albania. Rivista Italiana di Paleontologia e Stratigrafia, 105(2): 251-266 http://www.researchgate.net/publication/291841240_Conodont_biostratigraphy_of_Triassic_pelagic_strata_Albania |
| Mietto, P., Fratoni, R. P., Perri, M. C., 1991. Spathian and Aegean Conodonts from the Capelluzo Calcarenites of the Monte Facito Group. Memorie di Scienze Geologische, XLIII: 305-317 http://www.researchgate.net/publication/284937502_Spathian_and_aegean_conodonts_from_the_capelluzo_calcarenites_of_the_Monte_Facito_Group |
| Mosher, L. C., 1968. Triassic Conodonts from Western North America and Europe and Their Correlation. Journal of Paleontology, 42: 895-946 |
| Mosher, L. C., 1970. New Conodont Species as Triassic Guide Fossils. Journal of Paleontology, 44(4): 737-742 |
| Mosher, L. C., Clark, D. L., 1965. Middle Triassic Conodonts from the Prida Formation of Northwestern Nevada. Journal of Paleontology, 39: 551-565 |
| Müller, K. J., 1962. Zur Systematischen Einteilung der Conodontophorida. Paläontologische Zeitschrift, 36(1): 109-117. https://doi.org/10.1007/bf02989634 |
| Narkiewicz, K., 1999. Conodont Biostratigraphy of the Muschelkalk (Middle Triassic) in the Central Part of the Polish Lowlands. Geological Quarterly, 43(3): 313-328 http://www.researchgate.net/publication/269989645_Conodont_biostratigraphy_of_the_Muschelkalk_Middle_Triassic_in_the_central_part_of_the_Polish_Lowlands |
| Nicora, A., 1977. Lower Anisian Platform-Conodonts from the Tethys and Nevada: Taxonomic and Stratigraphic Revision. Palaeontographica Abteilung A, 157(1-3): 88-107 http://www.schweizerbart.de/papers/pala/detail/A157/70922/Lower_Anisian_Platform_Conodonts_from_the_Tethys_a |
| Nogami, Y., 1968. Trias-Conodonten von Timor, Malaysien und Japan (Palaeontological Study of Portuguese Timor, 5). Memoirs of the Faculty of Science, Kyoto University, Series of Geology and Mineralogy, 34: 115-136 http://ci.nii.ac.jp/naid/120005466484/ |
| Orchard, M. J., 1991a. Late Triassic Conodont Biochronology and Biostratigraphy of the Kunga Group, Queen Charlotte Islands, British Columbia. In: Woodsworth, G. W., ed., Evolution and Hydrocarbon Potential of the Queen Charlotte Basin, British Columbia. Geological Survey of Canada Paper, 1990-10: 173-193. https://doi.org/10.4095/131971 |
| Orchard, M. J., 1991b. Upper Triassic Conodont Biochronology and New Index Species from the Canadian Cordillera. In: Orchard, M. J., McCracken, A. D., eds., Ordovician to Triassic Conodont Paleontology of the Canadian Cordillera. Geological Survey of Canada, Bulletin, 417: 299-335. https://doi.org/10.4095/132442 |
| Orchard, M. J., 1994. Conodont Biochronology around the Early-Middle Triassic Boundary: New Data from North America, Oman and Timor. Memoires de Geologie (Lausanne), 22: 105-115 http://www.researchgate.net/publication/273693586_Conodont_biochronology_around_the_Early-Middle_Triassic_boundary_new_data_from_North_America_Oman_and_Timor |
| Orchard, M. J., 2005. Multielement Conodont Apparatuses of Triassic Gondolelloidea. Special Papers in Palaeontology, 73(73): 73-101 http://www.researchgate.net/publication/273694027_Multielement_Conodont_Apparatuses_of_Triassic_Gondolelloidea |
| Orchard, M. J., 2007. New Conodonts and Zonation, Ladinian-Carnian Boundary Beds, British Columbia, Canada. In: Lucas, S. G., Spielmann, J. A., eds., The Global Triassic. New Mexico Museum of Natural History and Science Bulletin, 41: 321-330 |
| Orchard, M. J., 2013. Five New Genera of Conodonts from the Carnian-Norian Boundary Beds of Black Bear Ridge, Northeast British Columbia, Canada. In: Tanner, L. H., Spielmann, J. A., Lucas, S. G., eds., The Triassic System. New Mexico Museum of Natural History and Science Bulletin, 61: 445-437 |
| Orchard, M. J., 2014. Conodonts from the Carnian-Norian Boundary (Upper Triassic) of Black Bear Ridge, Northeasern British Columbia, Canada. New Mexico Museum of Natural History and Science Bulletin, 64: 139 http://www.researchgate.net/publication/292234849_Conodonts_from_the_Carnian-Norian_boundary_Upper_Triassic_of_Black_Bear_Ridge_Northeasern_British_Columbia_Canada |
| Orchard, M. J., Cordey, F., Rui, L., et al., 2001. Biostratigraphic and Biogeographic Constraints on the Carboniferous to Jurassic Cache Creek Terrane in Central British Columbia. Canadian Journal of Earth Sciences, 38(4): 551-578. https://doi.org/10.1139/e00-120 |
| Orchard, M. J., Lehrmann, D. J., Wei, J. Y., et al., 2007a. Conodonts from the Olenekian-Anisian Boundary Beds, Guandao, Guizhou Province, China. New Mexico Museum of Natural History and Science Bulletin, 41: 347-354 http://www.researchgate.net/publication/251555588_Conodonts_from_the_Olenekian-Anisian_boundary_beds_Guandao_Guizhou_Province_China |
| Orchard, M. J., Gradinăru, E., Nicora, A., 2007b. A Summary of the Conodont Succession around the Olenekian-Anisian Boundary at Deşli Caira, North Dobrogea, Romania. New Mexico Museum of Natural History and Science Bulletin, 41: 341-346 |
| Orchard, M. J., Rieber, H., 1999. Multielement Neogondolella (Conodonta, Upper Permian-Middle Triassic). In: Serpagli, E., ed., Studies on conodonts. Proceedings of the Seventh European Conodont Symposium. June 1998, Bologna Modena, Italy. Bollettino della Societa Palaeontologica Italiana, 37: 475-488 |
| Pander, C. H., 1856. Monographie der Fossilen Fische des Silurischen Systems der Russisch-Baltischen Gouvernements. Buchdruckerei der Kaiserlichen Akademie der Wissenschaften, St. Petersburg. 91 |
| Pisa, G., Perri, C., Veneri, P., 1980. Upper Anisian Conodonts from Dont and M. Bivera Formations, Southern Alps (Italy). Rivista Italiana di Paleontologia e Stratigrafia, 85: 807-828 http://www.researchgate.net/publication/281135966_Upper_Anisian_conodonts_from_Dont_and_M_Bivera_Formations_southern_Alps_Italy |
| Rigo, M., Preto, N., Roghi, G., et al., 2007. A Rise in the Carbonate Compensation Depth of Western Tethys in the Carnian (Late Triassic): Deep-Water Evidence for the Carnian Pluvial Event. Palaeogeography, Palaeoclimatology, Palaeoecology, 246(2/3/4): 188-205. https://doi.org/10.1016/j.palaeo.2006.09.013 |
| Sudar, M. N., Budurov, K., 1979. New Conodonts from the Triassic in Yugoslavia and Bulgaria. Geologica Balcanica, 9(3): 47-52 http://www.researchgate.net/publication/284978298_New_conodonts_from_the_Triassic_in_Yugoslavia_and_Bulgaria |
| Sun, Z. Y., Sun, Y. L., Hao, W. C., et al., 2006. Conodont Evidence for the Age of the Panxian fauna, Guizhou, China. Acta Geologica Sinica, 80(5): 621-630 http://www.cqvip.com/Main/Detail.aspx?id=23139794 |
| Sun, Z. Y., Hao, W. -C., Sun, Y. -L., et al., 2009. The Conodont Genus Nicoraella and a New Species from the Anisian of Guizhou, South China. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 252(2): 227-235. https://doi.org/10.1127/0077-7749/2009/0252-0227 |
| Sweet, W. C., 1970. Uppermost Permian and Lower Triassic Conodonts of the Salt Range and Trans-İndus Ranges, West Pakistan. In: Kummel, B., Teichert, C., eds., Stratigraphic Boundary Problems: Permian and Triassic of West Pakistan. Volume Special Publication 4. University Press of Kansas, Lawrence. 207-275 |
| Tatge, U., 1956. Conodonten aus dem Germanischen Muschelkalk. Paläontologische Zeitschrift, 30(3/4): 129-147. https://doi.org/10.1007/bf03041777 |
| Trammer, J., 1975. Stratigraphy and Facies Development of the Muschelkalk in the South-Western Holy Cross Mts. Acta Geologica Polonica, 25: 179-216 http://dialnet.unirioja.es/servlet/articulo?codigo=265509 |
| Tüysüz, O., Aksay, A., Yiğitbaş, E., 2004. Batı Karadeniz Bolgesi Litostratigrafi Birimleri. Turkiye Stratigrafi Komitesi Litostratigrafi Birimleri Serisi-1, General Directorate of Mineral Research and Exploration Publications, Ankara, Turkey. 92 (in Turkish) |
| Wang, Z. H., Dai, J. Y., 1981. Triassic Conodonts from the Jiangyou-Beichuan Area, Sichuan Province. Acta Micropalaeontologica Sinica, 20: 138-150. https://doi.org/10.19800/j.cnki.aps.1981.02.005 |
| Zawidzka, K., 1975. Conodont Stratigraphy and Sedimentary Environment of the Muschelkalk in Upper Silesia. Acta Geologica Polonica, 25(2): 217-257 http://www.researchgate.net/publication/286780854_Conodont_stratigraphy_and_sedimentary_environment_of_the_Muschelkalk_in_Upper_Silesia |
| Zhang, Q. Y., Zhou, C. Y., Lu, T., et al., 2009. A Conodont-Based Middle Triassic Age Assignment for the Luoping Biota of Yunnan, China. Science in China Series D: Earth Sciences, 52(10): 1673-1678. https://doi.org/10.1007/s11430-009-0114-z |

Figures(6)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: