| Citation: | Andrew Simpson, David Mathieson, Jiri Fryda, Barbora Frydová. Summary of East Gondwanan Conodont Data through the Ireviken Event at Boree Creek. Journal of Earth Science, 2021, 32(3): 512-523. doi: 10.1007/s12583-021-1310-9

|

| Adrian, J., 1971. Stratigraphic Units of the Molong District, New South Wales. Geological Survey of New South Wales, Records, 13(4): 179-198 |
| Armstrong, H. A., 1990. Conodonts from the Upper Ordovician-Lower Silurian Carbonate Platform of North Greenland. Bulletin, Grønlands Geologiske Undersøgelse, 159: 1-151 doi: 10.34194/bullggu.v159.6709 |
| Barrick, J. E., Klapper, G., 1976. Multielement Silurian (Late Llandoverian-Wenlockian) Conodonts of the Clarita Formation, Arbuckle Mountains, Oklahoma, and Phylogeny of Kockelella. Geologica et Palaeontologica, 10: 59-100 http://www.researchgate.net/publication/284762363_Multielement_Silurian_late_Llandoverian-Wenlockian_conodonts_of_the_Clarita_Formation_Arbuckle_Mountains_Oklahoma_and_phylogeny_of_Kockelella |
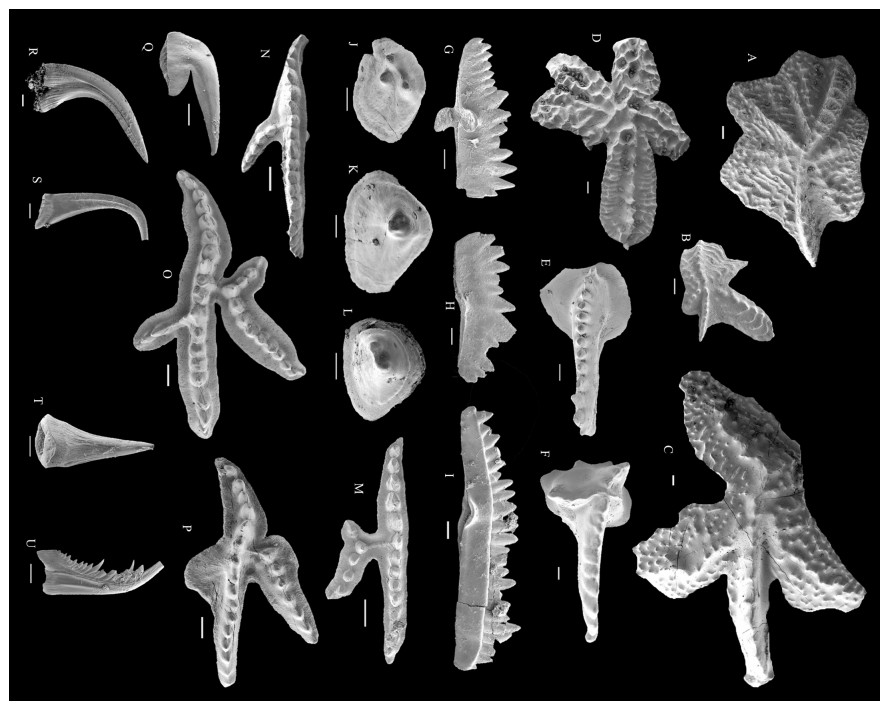
| Bischoff, G. C. O., 1997. Amelia mischa n. sp. (Conodonta) from Late Llandoverian and Early Wenlockian Strata of Midwestern New South Wales. Neues Jahrbuch für Geologie und Paläontologie-Monatshefte, 1997(8): 477-488. https://doi.org/10.1127/njgpm/1997/1997/477 http://www.researchgate.net/publication/293077966_Amelia_mischa_n_sp_Conodonta_from_late_Llandoverian_and_early_Wenlockian_strata_of_midwestern_New_South_Wales |
| Bischoff, G. C. O., 1998. New Species of Panderodus (Conodonta) from Late Llandoverian and Early Wenlockian Strata of Midwestern New South Wales. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 210(2): 267-288. https://doi.org/10.1127/njgpa/210/1998/267 |
| Bischoff, G. C. O., 1986. Early and Middle Silurian Conodonts from Midwestern New South Wales. Courier Forschungsinstitut Senckenberg, 89: 1-337 http://www.researchgate.net/publication/308340881_Early_and_Middle_Silurian_conodonts_from_midwestern_New_South_Wales |
| Brand, U., Azmy, K., Veizer, J., 2006. Evaluation of the Salinic I Tectonic, Cancañiri Glacial and Ireviken Biotic Events: Biochemostratigraphy of the Lower Silurian Succession in the Niagara Gorge Area, Canada and USA. Palaeogeography, Palaeoclimatology, Palaeoecology, 241(2): 192-213. https://doi.org/10.1016/j.palaeo.2006.03.004 |
| Branson, E. B., Mehl, M. G., 1933. Conodont Studies No. 1: Conodonts from the Bainbridge (Silurian) of Missouri. University of Missouri Studies, 8(1): 39-52 |
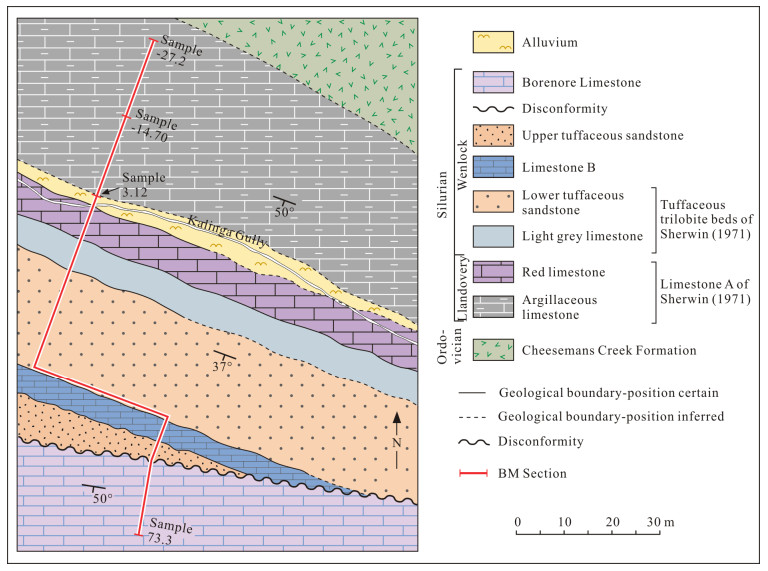
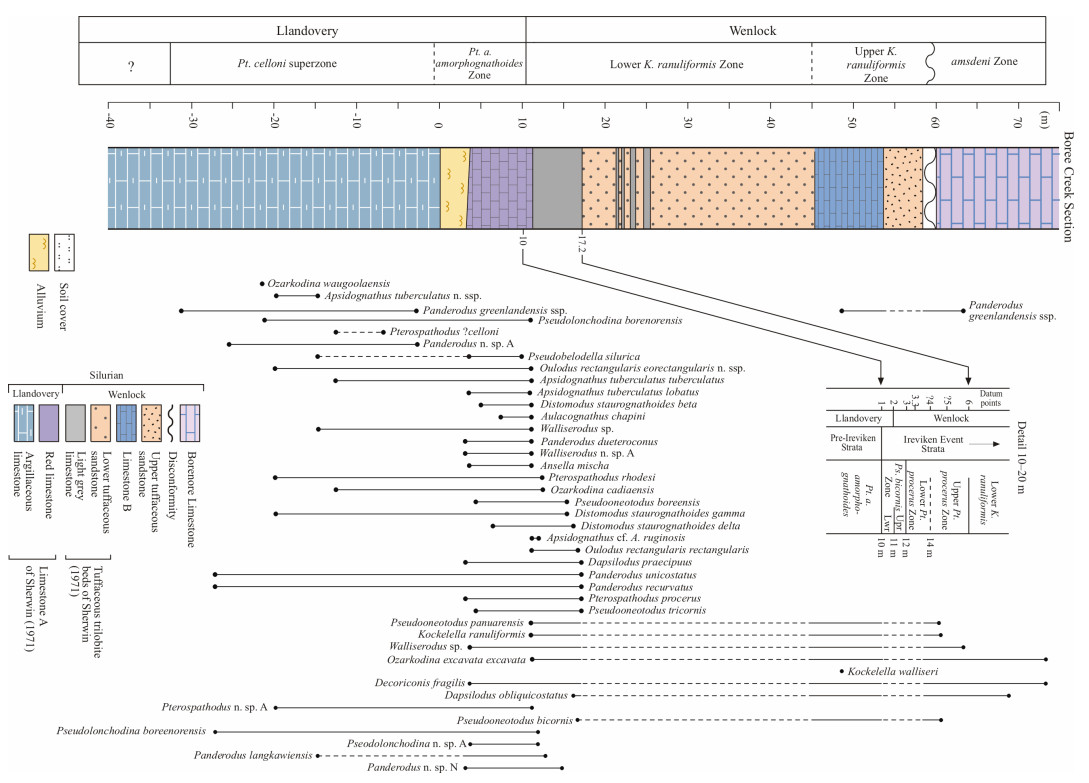
| Cockle, P., 1999. Conodont Data in Relation to Time Space and Environmental Relationships in the Siluran (Late Llandovery-Ludlow) Succession at Boree Creek (New South Wales, Australia). Abhandlungen der Geologischen Bundesanstalt, 54: 107-133 |
| Cramer, B. D., Saltzman, M. R., 2005. Sequestration of 12C in the Deep Ocean during the Early Wenlock (Silurian) Positive Carbon Isotope Excursion. Palaeogeography, Palaeoclimatology, Palaeoecology, 219(3/4): 333-349. https://doi.org/10.1016/j.palaeo.2005.01.009 |
| Downes, P. M., Colquhoun, G. P., Blevin, P. L., et al., 2013. Bathurst 1: 250 000 Metallogenic Map (2nd Edition), Geological Survey of New South Wales, Sydney |
| Drygant, D. M., 1974. Simple Conodonts from the Silurian and Lowermost Devonian of the Volyno-Podolia. Paleontologicheskii Sbornik, 10(2): 64-70 |
| Frýda, J., Lehnert, O., Joachimski, M., 2015. First Record of the Early Sheinwoodian Carbon Isotope Excursion (ESCIE) from the Barrandian Area of Northwestern Peri-Gondwana. Estonian Journal of Earth Sciences, 64(1): 42-46. https://doi.org/10.3176/earth.2015.08 |
| Frýda, J., Simpson, A. J., Frýdová, B., 2019. First Complete Record of the Early Sheinwoodian Carbon Isotope Anomaly from Australia. In: Petti, F. M., Innamorati, G., Carmina, B., eds., 3rd International Congress on Stratigraphy-Strati 2019: Abstract Book. Societa Geologica Italiana, Milan |
| Glen, R. A., Meffre, S., Scott, R. J., 2007. Benambran Orogeny in the Eastern Lachlan Orogen, Australia. Australian Journal of Earth Sciences, 54(2/3): 385-415. https://doi.org/10.1080/08120090601147019 |
| Helfrich, C. T., 1975. Silurian Conodonts from the Wills Mountain Anticline, Virginia, West Virginia, and Maryland. Geological Society of America, Special Paper, 161: 1-82 http://ci.nii.ac.jp/ncid/BA55077019 |
| Holloway, D. J., Lane, P. D., 1998. Effaced Styginid Trilobites from the Silurian of New South Wales. Palaeontology, 41(5): 853-896 |
| Igo, H., Koike, T., 1967. Ordovician and Silurian Conodonts from the Langkawi Islands, Malaya, Part 1. Geology and Palaeontology of Southeast Asia. In: Kobayashi, T., Toriyama, R., eds., Geology and Palaeontology of Southeast Asia 3. University of Tokyo Press, Tokyo. 3: 1-35 |
| Jell, J. S., Talent, J. A., 1989. Australia: The Most Instructive Sections. In: Holland, C. H., Bassett, M. G., eds., A Global Standard for the Silurian System. National Museum of Wales, Geological Series, 9: 183-200 |
| Jenkins, C. J., 1978. Llandovery and Wenlock Stratigraphy of the Panuara Area, Central New South Wales. Proceedings of the Linnean Society of New South Wales, 102(3): 109-130 |
| Jeppsson, L., 1990. An Oceanic Model for Lithological and Faunal Changes Tested on the Silurian Record. Journal of the Geological Society, 147(4): 663-674. https://doi.org/10.1144/gsjgs.147.4.0663 |
| Jeppsson, L., 1997a. The Anatomy of the Mid-Early Silurian Ireviken Event and a Scenario for P-S Events. In: Brett, C. E., Baird, G. C., eds., Paleontologic Events-Stratigraphic, Ecological and Evolutionary Implications. Columbia University Press, New York. 451-492 |
| Jeppsson, L., 1997b. A New Latest Telychian, Sheinwoodian and Early Homerian (Early Silurian) Standard Conodont Zonation. Transactions of the Royal Society of Edinburgh: Earth Sciences, 88(2): 91-114. https://doi.org/10.1017/s02635933000068544 doi: 10.1017/S0263593300006854 |
| Jeppsson, L., 1998. Silurian Oceanic Events, Summary of General Characteristics. In: Landing, E., Johnson, M. E., eds., Silurian Cycles. New York State Museum Bulletin, 491: 239-257 |
| Jeppsson, L., 1984. Sudden Appearances of Silurian Conodont Lineages: Provincialism or Special Biofacies?. Geological Society of America, 196: 103-112. https://doi.org/10.1130/spe196-p103 |
| Jeppsson, L., Männik, P., 1993. High-Resolution Correlations between Gotland and Estonia near the Base of the Wenlock. Terra Nova, 5(4): 348-358. https://doi.org/10.1111/j.1365-3121.1993.tb00268.x |
| Kaljo, D., Grytsenko, V., Martma, T., et al., 2007. Three Global Carbon Isotope Shifts in the Silurian of Podolia (Ukraine): Stratigraphical Implications. Estonian Journal of Earth Sciences, 56(4): 205-220. https://doi.org/10.3176/earth.2007.02 |
| Kaljo, D., Kiipli, T., Martma, T., 1997. Carbon Isotope Event Markers through the Wenlock-Pridoli Sequence at Ohesaare (Estonia) and Priekule (Latvia). Palaeogeography, Palaeoclimatology, Palaeoecology, 132(1/2/3/4): 211-223. https://doi.org/10.1016/S0031-0182(97)00065-5 |
| Loydell, D. K., 1998. Early Silurian Sea-Level Changes. Geological Magazine, 135(4): 447-471. https://doi.org/10.1017/s0016756898008917 doi: 10.1017/S0016756898008917 |
| Loydell, D. K., Frýda, J., 2007. Carbon Isotope Stratigraphy of the Upper Telychian and Lower Sheinwoodian (Llandovery-Wenlock, Silurian) of the Banwy River Section, Wales. Geological Magazine, 144(6): 1015-1019. https://doi.org/10.1017/s0016756807003895 doi: 10.1017/S0016756807003895 |
| Männik, P., 2007. An Updated Telychian (Late Llandovery, Silurian) Conodont Zonation Based on Baltic Faunas. Lethaia, 40(1): 45-60. https://doi.org/10.1111/j.1502-3931.2006.00005.x |
| Männik, P., 1998. Evolution and Taxonomy of the Silurian Conodont Pterospathodus. Palaeontology, 41(5): 1001-1050 |
| Männik, P., Aldridge, R. J., 1989. Evolution, Taxonomy and Relationships of the Silurian Conodont Pterospathodus. Palaeontology, 32(4): 893-906 |
| Molloy, P. D., 2006. Ananlysis of the Ireviken Extinction Event in the Boree Creek Formation, New South Wales, Australia based on Late Llandovery-Early Wenlock Conodonts: [Dissertation]. Macquarie University, Sydney |
| Molloy, P. D., Simpson, A., 2012. An Analysis of the Ireviken Event in the Boree Creek Formation, New South Wales, Australia. Earth and Life. Springer Netherlands, Dordrecht. 615-630 |
| Munnecke, A., Samtleben, C., Bickert, T., 2003. The Ireviken Event in the Lower Silurian of Gotland, Sweden-Relation to Similar Palaeozoic and Proterozoic Events. Palaeogeography, Palaeoclimatology, Palaeoecology, 195(1/2): 99-124. https://doi.org/10.1016/s0031-0182(03)00304-3 |
| Percival, I. G., Glen, R. A., 2007. Ordovician to Earliest Silurian History of the Macquarie Arc, Lachlan Orogen, New South Wales. Australian Journal of Earth Sciences, 54(2/3): 143-165. https://doi.org/10.1080/08120090601146789 |
| Percival, I. G., Webby, B. D., Pickett, J. W., 2001. Ordovician (Bendigonian, Darriwilian to Gisbornian) Faunas from the Northern Molong Volcanic Belt of Central New South Wales. Alcheringa: An Australasian Journal of Palaeontology, 25(2): 211-250. https://doi.org/10.1080/03115510108619105 |
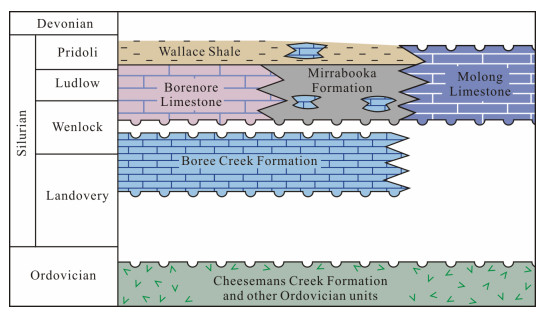
| Pickett, J., 1982. The Silurian System in New South Wales. Geological Survey of New South Wales, Bulletin, 29: 1-264 http://www.researchgate.net/publication/259297653_Pickett_JW_editor_1982a_The_Silurian_System_in_New_South_Wales_Bulletin_of_the_Geological_Survey_of_NSW_29_1-264 |
| Pogson, D. J., Watkins, J. J., 1998. Bathurst 1: 250 000 Geological Sheet. SI/55-8: Explanatory Notes. Australian Geological Survey Organisation and Geological Survey of New South Wales, SI/55-8 |
| Racki, G., Baliński, A., Wrona, R., et al., 2012. Faunal Dynamics across the Silurian-Devonian Positive Isotope Excursions (δ13C, δ18O) in Podolia, Ukraine: Comparative Analysis of the Ireviken and Klonk Events. Acta Palaeontologica Polonica, 57(4): 795-832. https://doi.org/10.4202/app.2011.0206 |
| Saltzman, M. R., 2001. Silurian δ13C Stratigraphy: A View from North America. Geology, 29(8): 671-674. https://doi.org/10.1130/0091-7613(2001)0290671:scsavf>2.0.co;2 doi: 10.1130/0091-7613(2001)029<0671:SCSAVF>2.0.CO;2 |
| Savage, N. M., 1985. Silurian (Llandovery-Wenlock) Conodonts from the Base of the Heceta Limestone, Southeastern Alaska. Canadian Journal of Earth Sciences, 22: 711-727 doi: 10.1139/e85-077 |
| Serpagli, E., Corradini, C., 1999. Taxonomy and Evolution of Kokellela (Conodonta) from the Silurian of Sardinia (Italy). Bollettino delle Societẚ Paleontologica Italiana, 37 (2/3): 275-289 |
| Sherwin, L., 1971. Stratigraphy of the Cheesemans Creek District, New South Wales. Geological Survey of New South Wales, Records, 13(4): 199-237 |
| Sherwin, L., Rickards, B., 2002. Late Silurian (Pridoli) Graptolites from the Wallace Shale, New South Wales. Alcheringa: An Australasian Journal of Palaeontology, 26(1): 87-101. https://doi.org/10.1080/03115510208619245 |
| Simpson, A., 1995. Silurian Conodont Biostratigraphy in Australia: A Review and Critique. Courier Forschungsinstitut Senckenberg, 182: 325-345 |
| Simpson, A., 1999. Early Silurian Conodonts from the Quinton Formation, Northeastern Australia. Abhandlungen der Geologisches Bundesanstalt, 54: 181-199 |
| Simpson, A., Talent, J. A., 1996. Middle Palaeozoic Orogenic Events in the Southern Lachlan Fold Belt: Age Constraints from Conodont Data. Geological Society of Australia, Abstracts, 41: 396 |
| Simpson, A., Talent, J. A., 1995. Silurian Conodonts from the Headwaters of the Indi (Upper Murray) and Buchan Rivers, Southeastern Australia, and Their Implications. Courier Forschungsinstitut Senckenberg, 182: 79-215 |
| Sloan, T., Talent, J. A., Mawson, R., et al., 1995. Conodont Data from Silurian-Middle Devonian Carbonate Fans, Debris Flows, Allochthonous Blocks and Adjacent Autochthonous Platform Margins: Broken River and Camel Creek Areas, North Queensland, Australia. Courier Forschungsinstitut Senckenberg, 182: 1-78 |
| Talent, J. A., Berry, W. B. N., Boucot, A. J., et al., 1975. Correlation of the Silurian Rocks of Australia, New Zealand, and New Guinea. Geological Society of America Special Papers. Geological Society of America. 1-115. https://doi.org/10.1130/spe150-p1 |
| Talent, J. A., Mawson, R., Simpson, A., 2003a. Silurian of Australia and New Guinea. In: Landing, E., Johnson, M. E., eds., Silurian Lands and Seas-Paleogeography outside Laurentia. New York State Museum Bulletin, 493: 181-200 |
| Talent, J. A., Mawson, R., Simpson, A., 2003b. The "Lost" Early Ordovician-Devonian Georgetown Carbonate Platform of Northeastern Australia. Courier Forschungsinstitut Senckenberg, 242: 71-79 |
| Talent, J. A., Mawson, R., Andrew, A. S., et al., 1993. Middle Palaeozoic Extinction Events: Faunal and Isotopic Data. Palaeogeography, Palaeoclimatology, Palaeoecology, 104(1/2/3/4): 139-152. https://doi.org/10.1016/0031-0182(93)90126-4 http://www.sciencedirect.com/science/article/pii/0031018293901264 |
| Talent, J. A., Mawson, R., Simpson, A. J., et al., 2002. Palaeozoics of North East Queensland: Broken River Region. Post-5 Field Excursion Guidebook, IPC2002, July 11-17, 2002. Macquarie University Centre for Ecostratigraphy and Palaeobiology, Special Publication, 1: 1-82 http://www.researchonline.mq.edu.au/vital/access/manager/Repository/mq:12829 |
| Talent, J. A., Mawson, R., 1999. North-Eastern Molong Arch and Adjacent Hill End Trough (Eastern Australia): Mid-Palaeozoic Conodont Data and Implications. Abhandlungen der Geologischen Bundesanstalt, 54: 49-105 |
| Trotter, J. A., Williams, I. S., Barnes, C. R., et al., 2016. New Conodont δ18O Records of Silurian Climate Change: Implications for Environmental and Biological Events. Palaeogeography, Palaeoclimatology, Palaeoecology, 443: 34-48. https://doi.org/10.1016/j.palaeo.2015.11.011 |
| Valentine, J. L., Brock, G. A., Molloy, P. D., 2003. Linguliformean Brachiopod Faunal Turnover across the Ireviken Event (Silurian) at Boree Creek, Central-Western New South Wales, Australia. Courier Forschungsinstitut Senckenberg, 242: 301-328 |
| Vandenberg, A. H. M., 1998. The Benambran Deformation, Eastern Lachlan Fold Belt: Some Observations on a Complex Event. Geological Society of Australia, Abstracts, 49: 449 |
| Vecoli, M., Riboulleau, A., Versteegh, G. J. M., 2009. Palynology, Organic Geochemistry and Carbon Isotope Analysis of a Latest Ordovician through Silurian Clastic Succession from Borehole Tt1, Ghadamis Basin, Southern Tunisia, North Africa: Palaeoenvironmental Interpretation. Palaeogeography, Palaeoclimatology, Palaeoecology, 273(3/4): 378-394. https://doi.org/10.1016/j.palaeo.2008.05.015 |
| Walker, W. B., 1959. Palaeozoic Stratigraphy of the Area to the West of Borenore, NSW. Journal and Proceedings of the Royal Society of New South Wales, 93(1-4): 39-46 |
| Walliser, O., 1964. Conodonten des Silurs. Abhandlungen der Hessischen Landesamtes für Bodenforschung, 41: 1-106 |
| Wenzel, B., 1997. Isotopenstratigraphische Untersuchungen an Silurischen Abfolgen Und Deren Paläozeanographische Interpretation. Erlanger Geologische Abhandlungen, 129: 1-117 |
| Wenzel, B., Joachimski, M. M., 1996. Carbon and Oxygen Isotopic Composition of Silurian Brachiopods (Gotland/Sweden): Palaeoceanographic Implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 122(1/2/3/4): 143-166. https://doi.org/10.1016/0031-0182(95)00094-1 |
Figures(5)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: