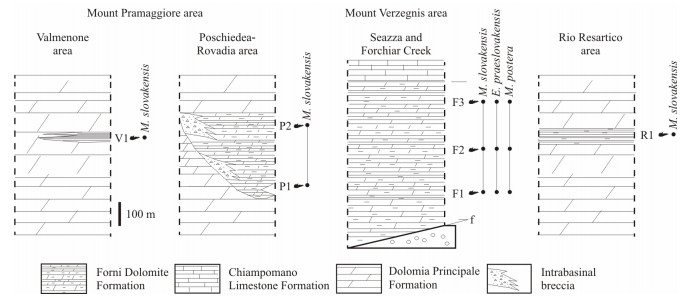
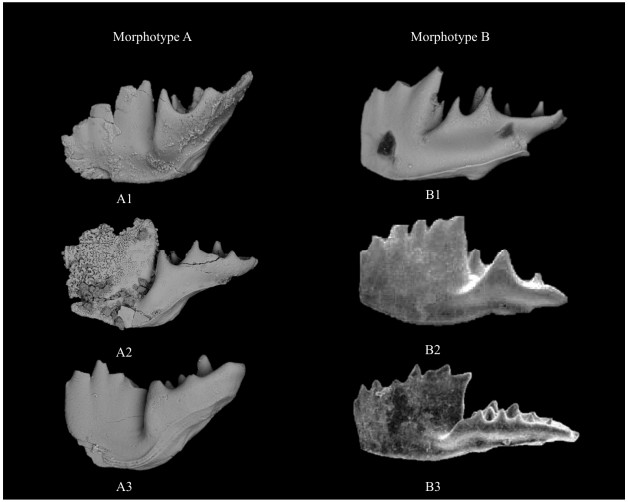
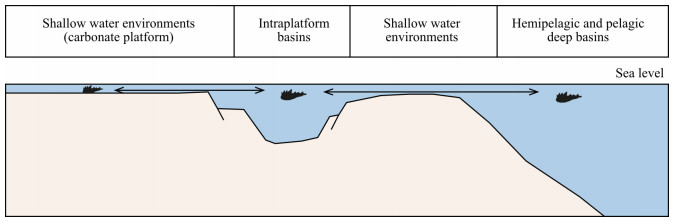
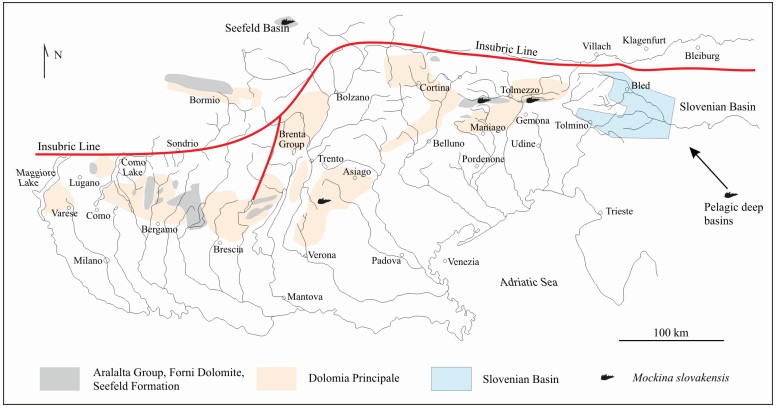
| Citation: | Yixing Du, Viktor Karádi, Guido Roghi, Maurizio Ponton, Andrea Cozzi, Manuel Rigo. Revision of the Conodont Mockina slovakensis and Its Paleogeographic Implications for the Upper Triassic Intraplatform Basins of the Alps. Journal of Earth Science, 2021, 32(3): 657-666. doi: 10.1007/s12583-021-1411-5

|

| Amodeo, F., Molisso, F., Kozur, H., et al., 1993. Age of Transitional Beds from «Cherty Limestones» (Calcari con Selce) to «Radiolarites» (Scisti Silicei) in the Lagonegro Domain (Southern Italy). First Evicence of Rhaetain Conodonts in Peninsular Italy. Bollettino del Servizio Geologico d'Italia, CX(3-22): 3-21 http://www.researchgate.net/publication/256484564_Age_of_Transitional_Beds_from_Cherty_Limestones_calcari_con_selce_to_Radiolarites_scisti_silicei_in_the_Lagonegro_Domain_Southern_Italy_First_evidence_of_Rhaetian_conodonts_in_peninsular_Italy |
| Balini, M., Bertinelli, A., Di Stefano, P., et al., 2010. The Late Carnian-Rhaetian Succession at Pizzo Mondello (Sicani Mountains). Albertiana, 39: 36-57 http://www.researchgate.net/publication/260640296_The_Late_Carnian-Rhaetian_succession_at_Pizzo_Mondello_Sicani_Mountains |
| Bazzucchi, P., Bertinelli, A., Ciarapica, G., et al., 2005. The Late Triassic-Jurassic Stratigraphic Succession of Pignola (Lagonegro-Molise Basin, Southern Apennines, Italy). Bollettino della Società Geologica Italiana, 124(1): 143-153 http://italianjgeo.geoscienceworld.org/content/124/1/143.abstract |
| Belvedere, M., Avanzini, M., Mietto, P., et al., 2008. Norian Dinosaur Footprints from the "Strada delle Gallerie" (Monte Pasubio, NE Italy). Studi Trentini Di Scienza Naturali, Acta Geologica, 83: 267-276 |
| Berra, F., 1995. Stratigraphic Evolution of a Norian Intraplatform Basin Recorded in the Quattervals Nappe (Austroalpine, Northern Italy) and Paleogeographic Implications. Eclogae Geologicae Helvetiae, 88(3): 501-528 http://www.researchgate.net/publication/257411320_Stratigraphic_evolution_of_a_Norian_intraplatform_basin_recorded_in_the_Quattervals_Nappe_(Austroalpine_Northern_Italy)_and_paleogeographic_implications |
| Berra, F., Jadoul, F., 1999. Stratigraphy, Paleogeography and Tectonic Setting of the Norian Succession of the Ortles Nappe (Central Austroalpine, Lombardy, Northern Italy). In: Gosso, G., Jadoul, F., Sella, M., et al., eds., 3rd Workshop on Alpine Geologicak Studies. Sept. 29-Oct. 1, 1997, Biella-Oropa. 78-89 |
| Berra, F., Delfrati, L., Ponton, M., 2007. Dolomia Principale. Carta Geologica d'Italia 1: 50 000. Catalogo Delle Formazioni-unità Tradizionali, (2): 63-72 |
| Berra, F., Jadoul, F., Anelli, A., 2010. Environmental Control on the End of the Dolomia Principale/Hauptdolomit Depositional System in the Central Alps: Coupling Sea-Level and Climate Changes. Palaeogeography, Palaeoclimatology, Palaeoecology, 290(1-4): 138-150. https://doi.org/10.1016/j.palaeo.2009.06.037 |
| Bertinelli, A., Casacci, M., Concheri, G., et al., 2016. The Norian/Rhaetian Boundary Interval at Pignola-Abriola Section (Southern Apennines, Italy) as a GSSP Candidate for the Rhaetian Stage: An Update. Albertiana, 43: 5-18 http://www.researchgate.net/publication/301673795_The_NorianRhaetian_boundary_interval_at_Pignola-Abriola_section_Southern_Apennines_Italy_as_a_GSSP_candidate_for_the_Rhaetian_Stage_an_update |
| Brandner, R., Poleschinski, W., 1986. Stratigraphie und Tektonik am Kalkalpensüdrand zwischen Zirl und Seefeld in Tirol (Exkursion D am 3. April 1986). Jahresberichte und Mitteilungen des Oberrheinischen Geologischen Vereins, 67-92 http://www.researchgate.net/publication/309050639_Stratigraphie_und_Tektonik_am_Kalkalpensudrand_zwischen_Zirl_und_Seefeld_in_Tirol_Exkursion_D_am_3_April_1986 |
| Budai, T., Kovács, S., 1986. Contributions to the Stratigraphy of the Rezi Dolomite Formation Metapolygnathus slovakensis (Conodonta, Upper Triassic) from the Keszthely Mts (W Hungary). Annual Report of the Geological Institute of Hungary, 1984: 175-191 |
| Budurov, K. J., Sudar, M. N., 1990. Late Triassic Conodont Stratigraphy. Courier Forschungsinstitut Senckenberg, 118: 203-239 |
| Buser, S., 1989. Development of the Dinaric and the Julian Carbonate Platforms and of the Intermediate Slovenian Basin (NW Yugoslavia). Bollettino della Società Geologica Italiana, 40: 313-320 http://www.researchgate.net/publication/285475171_Development_of_the_Dinaric_and_the_Julian_carbonate_platforms_and_of_the_intermediate_Slovenian_Basin_NW_Yugoslavia |
| Buser, S., 1996. Geology of Western Slovenia and Its Paleogeographic Evolution. In: Drobne, K., Goričan, Š., Kotnik, B., eds., The Role of Impact Processes and Biological Evolution of Planet Earth-International Workshop. ZRC Publ., ZRC SAZU, Ljubljana. 111-123 |
| Carulli, G. B., Longo Salvador, G., Ponton, M., et al., 1997. La Dolomia di Forni: Evoluzione di un Bacino Euxinico Tardo-Triassico nelle Prealpi Carniche. Bollettino della Società Geologica Italiana, 116(1): 95-107 |
| Carulli, G. B., Cozzi, A., Longo Salvador, G., et al., 2000. Geologia delle Prealpi Carniche (con Carta Geologica delle Prealpi Carniche alla Scala 1: 50 000 e Note Illustrative alla Carta Geologica delle Prealpi Carniche). Pubbl. Mus. Friul. St. Nat., Udine. 44 |
| Channell, J. E. T., Kozur, H. W., Sievers, T., et al., 2003. Carnian-Norian Biomagnetostratigraphy at Silická Brezová (Slovakia): Correlation to Other Tethyan Sections and to the Newark Basin. Palaeogeography, Palaeoclimatology, Palaeoecology, 191(2): 65-109. https://doi.org/10.1016/s0031-0182(02)006545 |
| Cousin, M., 1973. Le Sillon Slovene: Les Formations Triasiques, Jurassiques et Ndocomiennes au Nord-Est de Tohnin (Slovdnie Occidentale, Alpes Mdridionales) et leurs Affinite Dinarique. Bulletin de la Société Géologique de France, 15: 326-339 http://bsgf.geoscienceworld.org/content/S7-XV/3-4/326 |
| Cousin, M., 1981. Les Rapports Alpes-Dinarides: Les Confins de l'Italie et de Yougoslavie. Vol. I. Société Géologique du Nord, 5: 1-521 http://www.researchgate.net/publication/285798955_Les_rapports_Alpes-Dinarides |
| Cozzi, A., 2000. The Effects of Extensional Tectonics on Shallow Water Carbonate Platform Facies and Geometry: The Case of the Upper Triassic Dolomia Principale, Carnian Prealps, Northern Italy. AAPG Annual Convention, New Orleans, Louisiana. #90914 |
| Cozzi, A., 2002. Facies Patterns of a Tectonically-Controlled Upper Triassic Platform-Slope Carbonate Depositional System (Carnian Prealps, Northeastern Italy). Facies, 47(1): 151-178. https://doi.org/10.1007/bf02667711 |
| Cozzi, A., Hardie, L. A., 2003. Third-Order Depositional Sequences Controlled by Synsedimentary Extensional Tectonics: Evidence from Upper Triassic Carbonates of the Carnian Prealps (NE Italy). Terra Nova, 15(1): 40-45. https://doi.org/10.1046/j.1365-3121.2003.00461.x |
| Cozzi, A., Jäger, H., 2000. Sedimentology, Palynofacies and Significance of an Intraplatform Anoxic Depression within the Upper Triassic Dolomia Principale (Carnian Prealps, Northern Italy). In: Abstr. 80th Summer Meeting Società Geologica Italiana. Sept. 6-8, 2000, Trieste. 164-165 |
| Csillag, G., Budai, T., Gyalog, L., 1995. Contribution to the Upper Triassic Geology of the Keszthely Mountains (Transdanubian Range), Western Hungary. Acta Geologica Hungarica, 38(2): 111-129 http://www.researchgate.net/publication/297794391_Contribution_to_the_Upper_Triassic_geology_of_the_Keszthely_Mountains_Transdanubian_Range_western_Hungary |
| Dal Corso, J., Ruffell, A., Preto, N., 2019. Carnian (Late Triassic) C-Isotope Excursions, Environmental Changes, and Biotic Turnover: A Global Perturbation of the Earth's Surface System. Journal of the Geological Society, 176(1): 129-131. https://doi.org/10.1144/jgs2018-217 |
| Dalla Vecchia, F. M., Muscio, G., 1991. I Fossili della Dolomia di Forni (Norico-Prealpi Carniche, Udine). Paleocronache, (1): 9-19 http://www.mendeley.com/research/i-fossili-della-dolomia-di-forni-norico-prealpi-carniche-udine/ |
| Donofrio, D. A., Brandner, R., Poleschinski, W., 2003. Conodonten der Seefeld-Formation: Ein Beitrag zur Bio-und Lithostratigraphie der Hauptdolomit-Plattform (Obertrias, Westliche Nördliche Kalkalpen, Tirol). Geologisch-Paläontologische Mitteilungen Innsbruck, 26: 91-107 http://www.researchgate.net/publication/242557101_Conodonten_der_Seefeld-Formation_Ein_Beitrag_zur_Bio-_und_Lithostratigraphie_der_Hauptdolomit-Plattform_Obertrias_Westliche_Nordliches_Kalkalpen_Tirol |
| Fantoni, R., Salvador, G. L., Podda, F., et al., 1998. L'unità a Laminiti Organiche nel Norico del Rio Resartico (Val Resia, Prealpi Giulie). Atti Ticinensi di Scienze Della Terra, 7(Suppl. ): 109-123 http://www.researchgate.net/publication/257935563_L'Unita_a_laminiti_organiche_nel_Norico_del_Rio_Resartico_Val_Resia_Prealpi_Giulie |
| Fruth, I., Scherreiks, R., 1984. Hauptdolomit-Sedimentary and Paleogeographic Models (Norian, Northern Calcareous Alps). Geologische Rundschau, 73(1): 305-319. https://doi.org/10.1007/bf01820372 |
| Giordano, N., Rigo, M., Ciarapica, G., et al., 2010. New Biostratigraphical Constraints for the Norian/Rhaetian Boundary: Data from Lagonegro Basin, Southern Apennines, Italy. Lethaia, 43: 573-586. https://doi.org/10.1111/j.1502-3931.2010.00219.x |
| Gullo, M., 1996. Conodont Biostratigraphy of Uppermost Triassic Deep-Water Calcilutites from Pizzo Mondello (Sicani Mountains): Evidence for Rhaetian Pelagites in Sicily. Palaeogeography, Palaeoclimate, Palaeoecology, 126: 309-323. https://doi.org/10.1016/s0031-0182(96)00043-0 |
| Haas, J., Kovács, S., Krystyn, L., et al., 1995. Significance of Late Permian-Triassic Facies Zones in Terrane Reconstructions in the Alpine-North Pannonian Domain. Tectonophysics, 242(1/2): 19-40. https://doi.org/10.1016/0040-1951(94)00157-5 |
| Haas, J., 2002. Origin and Evolution of Late Triassic Backplatform and Intraplatform Basins in the Transdanubian Range, Hungary. Geologica Carpathica, 53(3): 159-178 http://www.researchgate.net/publication/237277224_Origin_and_evolution_of_Late_Triassic_backplatform_and_intraplatform_basins_in_the_Transdanubian_Range_Hungary |
| Jadoul, F., 1985. Stratigrafia e Paleogeografia del Norico nelle Prealpi Bergamasche Occidentali. Rivista Italiana di Paleontologia e Stratigrafia, 91: 479-512 http://www.researchgate.net/publication/257483503_Stratigrafia_e_Paleogeografia_del_Norico_nelle_Prealpi_Bergamasche_Occidentali |
| Karádi, V., Cau, A., Mazza, M., et al., 2020. The Last Phase of Conodont Evolution during the Late Triassic: Integrating Biostratigraphic and Phylogenetic Approaches. Palaeogeography, Palaeoclimatology, Palaeoecology, 549: 109144. https://doi.org/10.1016/j.palaeo.2019.03.045 |
| Kaya, O., Helvaci, H., Kozur, H., et al., 2001. Late Norian Conodont Age for a Metacarbonate Unit in NW Anatolia, Turkey. Geobios, 34(5): 527-532. https://doi.org/10.1016/s0016-6995(01)80067-x |
| Kovács, S., Nagy, G., 1989. Contribution to the Age of the Avicula-and Halobia-Limestones (Feketehegy Limestone Formation) in Pilis Mts (NE Transdanubian Central Range, Hungary). Annual Report of the Hungarian Geological Institute, 1987: 95-129 http://www.researchgate.net/publication/288033455_Fekete-hegy_limestone_formation_in_the_pilis_mts_NE_transdanubian_central_range_Hungary |
| Kozur, H. W., 1972. Die Conodontengattung Metapolygnathus Hayashi 1968 und ihr Stratigraphischer wert. Geologisch-Paläontologische Mitteilungen Innsbruck, 2(11): 1-37 http://www.museocivico.rovereto.tn.it/patrimonio_abbonamenti_intermedio.jsp?area=69&ID_LINK=111098&page=2970&id_context=79137 |
| Martini, R., Zaninetti, L., Villeneuve, M., et al., 2000. Triassic Pelagic Deposits of Timor: Palaeogeographic and Sea-Level Implications. Palaeogeography, Palaeoclimatolology, Palaeoecology, 160(1/2): 123-151. https://doi.org/10.1016/s0031-0182(00)00062-6 |
| Mastandrea, A., Ietto, F., Neri, C., et al., 1997. Conodont Biostratigraphy of the Late Triassic Sequence of Monte Cocuzzo (Catena Costiera, Calabria, Italy). Rivista Italiana di Paleontologia e Stratigrafia, 103(2): 173-182 http://www.researchgate.net/publication/287678683_Conodont_biostratigraphy_of_the_late_triassic_sequence_of_Monte_Cocuzzo_Catena_Costiera_Calabria_Italy |
| Mastandrea, A., Perri, E., Neri, C., et al., 2003. Conodont Biostratigraphy of the Norian-Rhaetian Deposits in the Northern Calabria: The Valle Corvino and Grisolia Sections. Bollettino-Societa Paleontologica Italiana, 42(1/2): 39-48 http://www.researchgate.net/publication/288358127_Conodont_biostratigraphy_of_the_Norian-Rhaetian_deposits_in_the_Northern_Calabria_The_Valle_Corvino_and_Grisolia_sections |
| Mattavelli, L., Rizzini, A., 1974. Facies Euxiniche nelle Dolomie Noriche dell'Ampezzano (Udine): Petrografia e Sedimentologia. In: Contributi Strat. Paleog. Mesozoico Tetide. Memorie della Rivista Italiana di Paleontologia e Stratigrafia, 14: 111-140 |
| Mazza, M., Rigo, M., Gullo, M., 2012. Taxonomy and Biostratigraphic Record of the Upper Triassic Conodonts of the Pizzo Mondello Section (Western Sicily, Italy), GSSP Candidate for the Base of the Norian. Rivista Italiana di Paleontologia e Stratigrafia, 118(1): 85-130 http://www.researchgate.net/publication/290390666_Taxonomyand_biostratigraphic_record_of_the_uppertriassic_conodonts_of_the_pizzo_mondello_section_western_sicily_italy_gssp_candidate_for_the_base_of_the_norian |
| Meço, S., 1999. Conodont Biostratigraphy of Triassic Pelagic Strata, Albania. Rivista Italiana di Paleontologia e Stratigrafia, 105(2): 251-266 http://www.researchgate.net/publication/291841240_Conodont_biostratigraphy_of_Triassic_pelagic_strata_Albania |
| Moix, P., Kozur, H. W., Stampfli, G. M., et al., 2007. New Paleontological, Biostratigraphical and Paleogeographic Results from the Triassic of the Mersin Mélange, SE Turkey. The Global Triassic. New Mexico Museum of Natural History and Science Bulletin, 41: 282-311 http://www.researchgate.net/publication/286284961_New_paleontological_biostratigraphic_and_paleogeographic_results_from_the_Triassic_of_the_Mersin_Melange_SE_Turkey |
| Onoue, T., Tanaka, H., 2005. Late Triassic Bivalves from Sambosan Accretionary Complex, Southwest Japan, and Their Biogeographic Implications. Paleontological Research, 9(1): 15-25. https://doi.org/10.2517/prpsj.9.15 |
| Ponton, M., 2017. Dolomie Bituminose nella Dolomia Principale: La Miniera del Rio Resartico (Prealpi Giulie-Italia). Gortania: Geologia Paleontologia Paletnologia, 38: 9-37 (in Italian) |
| Rigo, M., Bertinelli, A., Concheri, G., et al., 2016. The Pignola-Abriola Section (Southern Apennines, Italy): A New GSSP Candidate for the Base of the Rhaetian Stage. Lethaia, 49: 287-306. https://doi.org/10.1111/let.12145 |
| Rigo, M., Mazza, M., Karádi, V., et al., 2018. New Upper Triassic Conodont Biozonation on of the Tethyan Realm. In: Tanner, L. H., ed., The Late Triassic World: Earth in a Time of Transition. Topics in Geobiology, 46: 189-235. https://doi.org/10.1007/978-3-319-68009-5_6 |
| Rigo, M., Preto, N., Roghi, G., et al., 2007. A Rise in the Carbonate Compensation Depth of Western Tethys in the Carnian (Late Triassic): Deep-Water Evidence for the Carnian Pluvial Event. Palaeogeography, Palaeoclimatology, Palaeoecology, 246(2-4): 188-205. https://doi.org/10.1016/j.palaeo.2006.09.013 |
| Rigo, M., Preto, N., Franceschi, M., et al., 2012. Stratigraphy of the Carnian-Norian Calcari con Selce Formation in the Lagonegro Basin, Southern Apennines. Rivista Italiana di Paleontologia e Stratigrafia, 118(1): 143-154 |
| Roghi, G., Mietto, P., Vecchia, D., 1995. Contribution to the Conodont Biostratigraphy of the Dolomia di Forni (Upper Triassic, Carnia, NE Italy). Memoire di Scienze Geologiche, 47: 125-133 http://www.researchgate.net/publication/256103845_Contribution_to_the_Conodont_Biostratigraphy_of_the_Dolomia_di_Forni_Upper_Triassic_Carnia_NE_Italy |
| Rožič, B., Kolar-Jurkovšek, T., Šmuc, A., 2009. Late Triassic Sedimentary Evolution of Slovenian Basin (Eastern Southern Alps): Description and Correlation of the Slatnik Formation. Facies, 55(1): 137-155. https://doi.org/10.1007/s10347-008-0164-2 |
| Schmid, S. M., Fügenschuh, B., Kissling, E., et al., 2004. Tectonic Map and Overall Architecture of the Alpine Orogen. Eclogae Geologicae Helvetiae, 97(1): 93-117. https://doi.org/10.1007/s00015-004-1113-x |
| Scotti, P., Fantoni, R., Podda, F., et al., 2002. Depositi Norici di Ambiente Anossico nelle Prealpi Friulane (Italia Nord-Orientale). Mem. Soc. Geol. It. , 57: 65-78 http://www.researchgate.net/publication/257935738_Depositi_norici_di_ambiente_anossico_nelle_Prealpi_Friulane_Italia_nord-_orientale?ev=prf_high |
| Simms, M. J., Ruffell, A. H., 1989. Synchroneity of Climatic Change and Extinctions in the Late Triassic. Geology, 17(3): 265-268. https://doi.org/10.1130/0091-7613(1989)017<0265:soccae>2.3.co;2 doi: 10.1130/0091-7613(1989)017<0265:soccae>2.3.co;2 |
| Trotter, J. A., Williams, I. S., Nicora, A., et al., 2015. Long-Term Cycles of Triassic Climate Change: A New δ18O Record from Conodont Apatite. Earth and Planetary Science Letters, 415:165-174. https://doi.org/10.1016/j.epsl.2015.01.038 |
| Yamashita, D., Kato, H., Onoue, T., et al., 2018. Integrated Upper Triassic Conodont and Radiolarian Biostratigraphies of the Panthalassa Ocean. Paleontological Research, 22(2): 167-197. https://doi.org/10.2517/2017pr020 |
Figures(6) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: