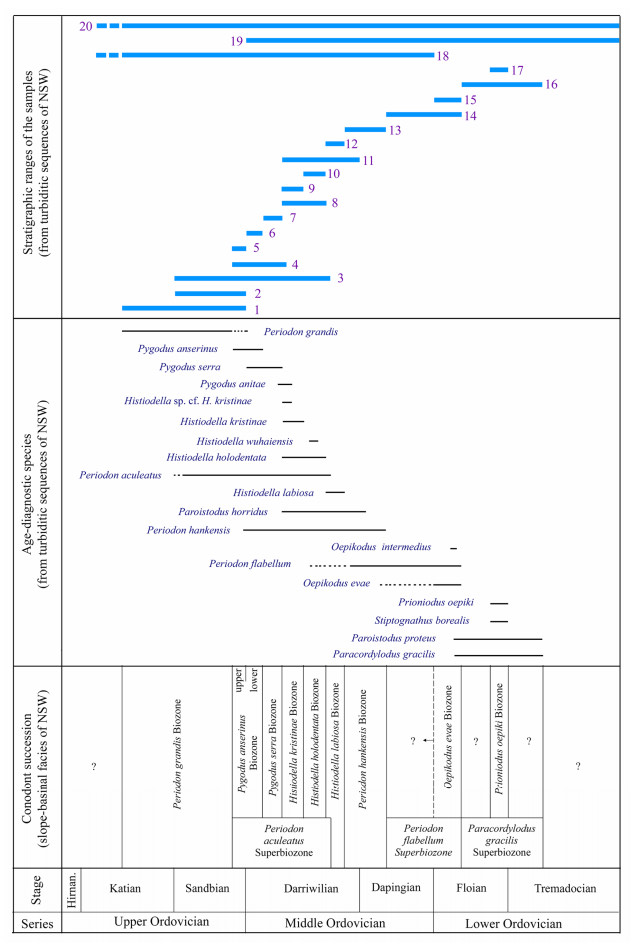
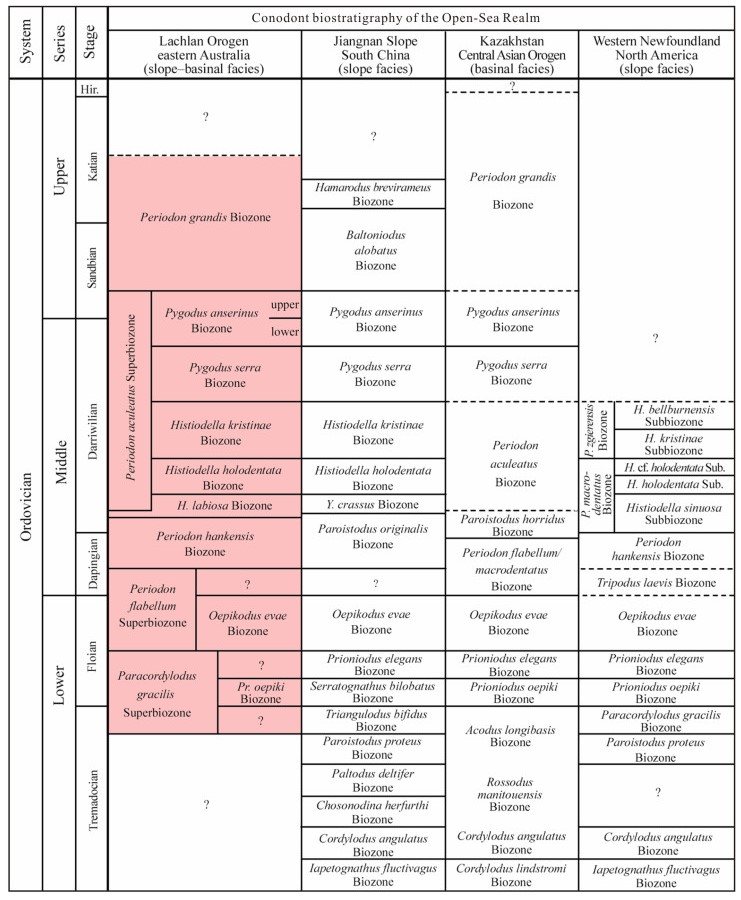
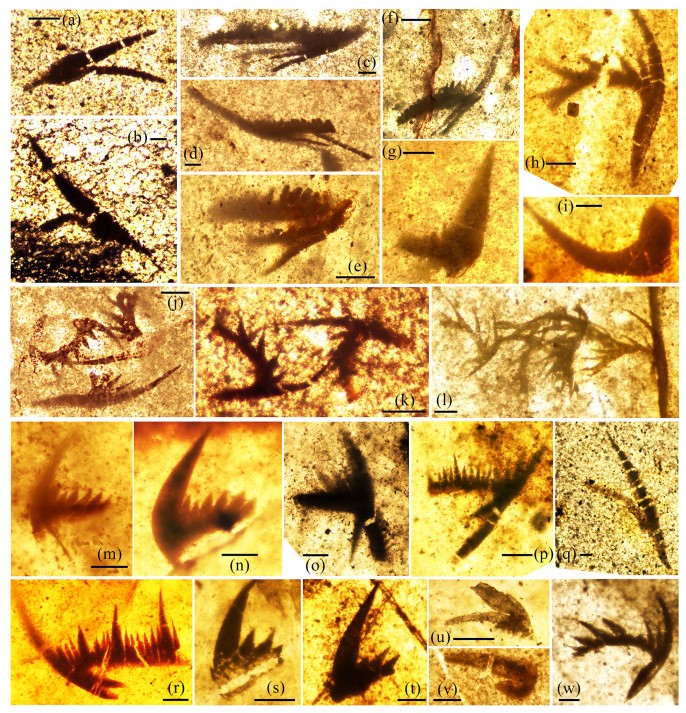
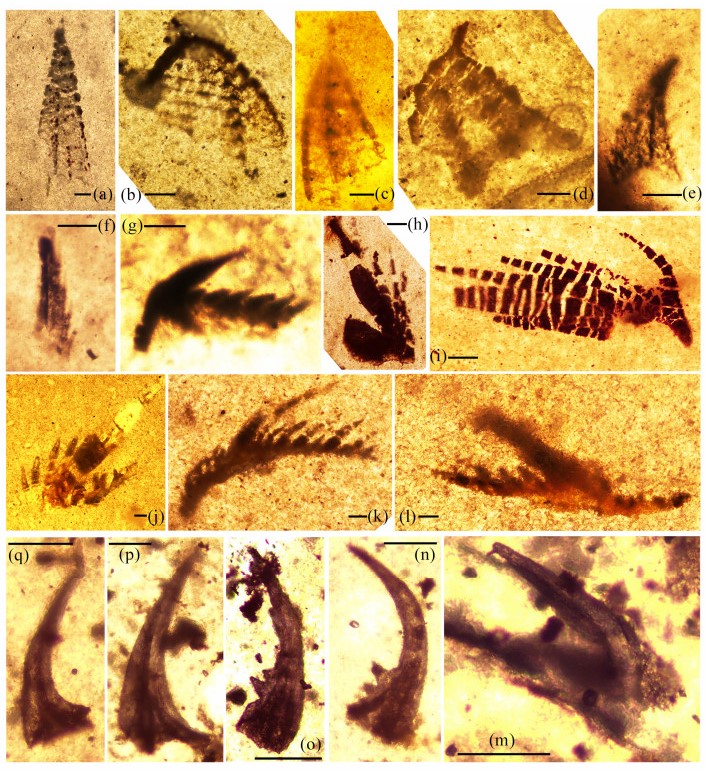
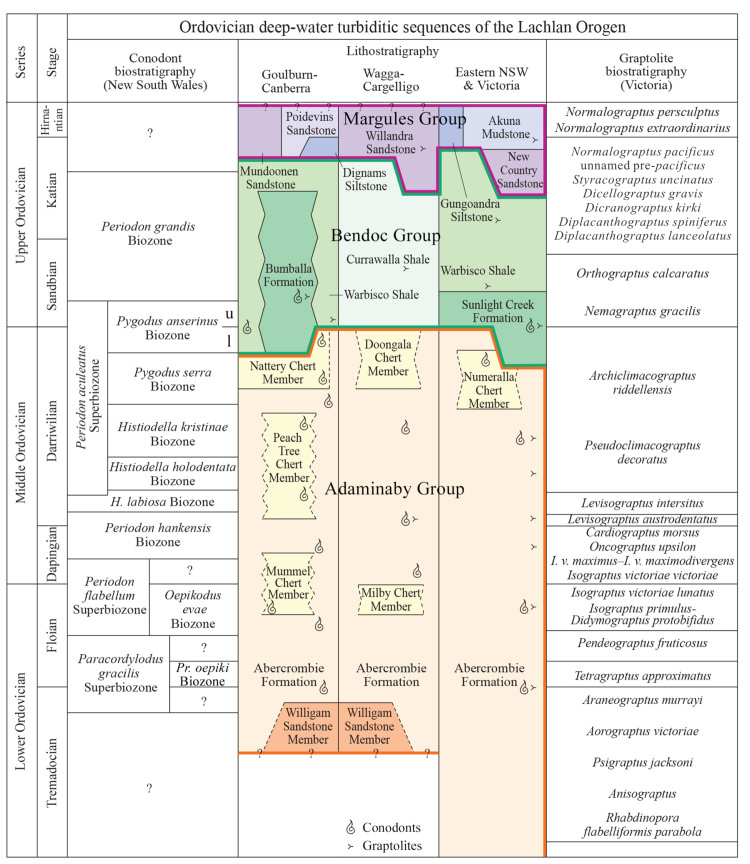
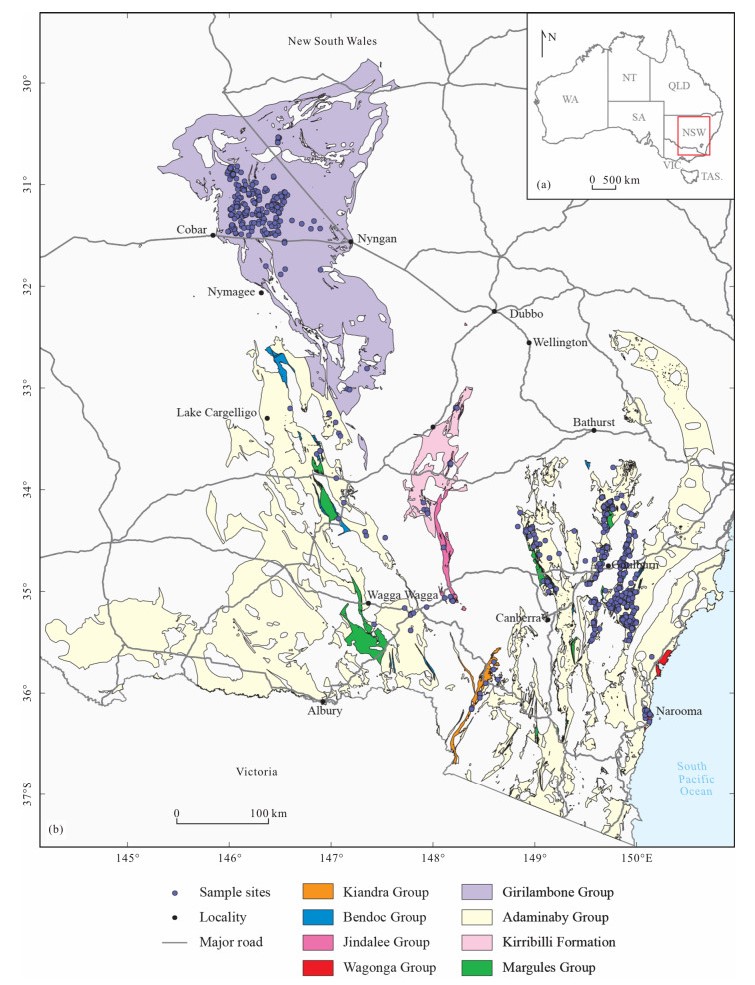
| Citation: | Yong Yi Zhen, Ian G. Percival, Phil Gilmore, Jodie Rutledge, Liann Deyssing. Conodont Biostratigraphy of Ordovician Deep-Water Turbiditic Sequences in Eastern Australia——A New Biozonal Scheme for the Open-Sea Realm. Journal of Earth Science, 2021, 32(3): 486-500. doi: 10.1007/s12583-021-1421-3

|

| An, T. X., 1981. Recent Progress in Cambrian and Ordovician Conodont Biostratigraphy of China. Geological Society of America Special Papers, 187: 209-226. https://doi.org/10.1130/spe187 http://www.researchgate.net/publication/278729976_Recent_Progress_in_Cambrian_and_Ordovician_Conodont_Biostratigraphy_of_China |
| Armstrong, H. A., Floyd, J. D., Barron, H. F., 2001. Conodont Biostratigraphy of the Crawford Group, Southern Uplands. British Geological Survey Research Report, RR/01/05. 1-36 http://www.researchgate.net/publication/255569571_Conodont_biostratigraphy_of_the_Crawford_Group_Southern_Uplands |
| Barnes, C. R., Poplawski, M. L. S., 1973. Lower and Middle Ordovician Conodonts from the Mystic Formation, Québec, Canada. Journal of Paleontology, 47(4): 760-790 |
| Bauer, J. A., 2010. Conodonts and Conodont Biostratigraphy of the Joins and Oil Creek Formations, Arbuckle Mountains, South-Central Oklahoma. Oklahoma Geological Survey Bulletin, 150: 1-44. http://ogs.ou.edu/docs/bulletins/b150.pdf http://www.researchgate.net/publication/266177220_conodonts_and_conodont_biostratigraphy_of_the_joins_and_oil_creek_formations_arbuckle_mountains_south-central_oklahoma |
| Bergström, S. M., 1983. Biogeography, Evolutionary Relationships, and Biostratigraphic Significance of Ordovician Platform Conodonts. Fossils and Strata, 15: 35-58 http://www.researchgate.net/publication/291977656_Biogeography_evolutionary_relationships_and_biostratigraphic_significance_of_Ordovician_platform_conodonts |
| Bruce, M. C., Percival, I. G., 2014. Geochemical Evidence for Provenance of Ordovician Cherts in Southeastern Australia. Australian Journal of Earth Sciences, 61(7): 927-950. https://doi.org/10.1080/08120099.2014.956792 |
| Colquhoun, G. P., Hughes, K. S., Deyssing, L., et al., 2020. New South Wales Seamless Geology Dataset, Version 2[Digital Dataset]. Geological Survey of New South Wales, Department of Regional NSW, Maitland |
| Cooper, B. J., 1981. Early Ordovician Conodonts from the Horn Valley Siltstone, Central Australia. Palaeontology, 24(1): 147-183 http://ci.nii.ac.jp/naid/10021382748 |
| Cooper, R. A., Nowlan, G. S., Williams, S. H., 2001. Global Stratotype Section and Point for Base of the Ordovician System. Episodes, 24(1): 19-28. https://doi.org/10.18814/epiiugs/2001/v24i1/005 |
| Deyssing, L., Trigg, S. J., Gilmore, P. J., et al., 2018. Old Friends Reunite: A New Proposal for an Ordovician Lachlan Supergroup in the Tasmanides. Geological Survey of NSW. Geological Survey Report No. GS 2018/1135 |
| Dubinina, S. V., 2000. Conodonts and Zonal Stratigraphy of the Cambrian-Ordovician Boundary Beds. Transactions of the Russian Academy of Sciences, Geological Institute, 517: 1-239 (in Russian) http://www.researchgate.net/publication/285976386_Conodonts_and_zonal_stratigraphy_of_the_Cambrian-Ordovician_boundary_deposits |
| Ethington, R. L., 1959. Conodonts of the Ordovician Galena Formation. Journal of Paleontology, 33: 257-292 |
| Ethington, R. L., Clark, D. L., 1982. Lower and Middle Ordovician Conodonts from the Ibex Area, Western Millard County, Utah. Brigham Young University Geology Studies, 28(2): 1-160 http://ci.nii.ac.jp/naid/10019474744 |
| Feltes, N. A., Albanesi, G. L., 2013. The Periodon and Paroistodus Conodont Biofacies in the Lower Member of the Las Aguaditas Formation (Middle Ordovician), Central Precordillera, Argentina. Conodonts from the Andes-The Third International Conodont Symposium, Mendoza, Argentina. Asociación Paleontológica Argentina Publicación Especial, 13: 17-23 http://www.researchgate.net/publication/303133824_The_Periodon_and_Paroistodus_conodont_biofacies_in_the_Lower_Member_of_the_Las_Aguaditas_Formation_Middle_Ordovician_central_Precordillera_Argentina |
| Glen, R. A., Stewart, I. R., Percival, I. G., 2004. The Narooma Terrane: Implications for the Construction of the Outboard Part of the Lachlan Orogen. Australian Journal of Earth Sciences, 51(6): 859-884. https://doi.org/10.1111/j.1400-0952.2004.01090.x |
| Graves, R. W., Ellison, S., 1941. Ordovician Conodonts of the Marathon Basin, Texas. University of Missouri, School of Mines and Metallurgy, Bulletin (Tech. Ser. ), 14: 1-26 http://ci.nii.ac.jp/naid/10021382754 |
| Hadding, A. R., 1913. Undre Dicellograptusskiffern I Skåne Jämte Några Dågra Därmed Dkvivalenta Bildningar. Lunds Universitets Årsskrift, Ny Följd, Afdelning 2, 9(15): 1-90 http://www.researchgate.net/publication/303272363_Undre_Dicellograptusskiffern_i_Skane_jamte_nagra_darmed_ekvivalenta_bildningar |
| Lamont, A., Lindstrom, M., 1957. Arenigian and Llandeilian Cherts Identified in the Southern Uplands of Scotland by Means of Conodonts, etc. Transactions of the Edinburgh Geological Society, 17(1): 60-70. https://doi.org/10.1144/transed.17.1.60 |
| Lindström, M., 1954. Conodonts from the Lowermost Ordovician Strata of South-Central Sweden. Geologiska Föreningen i Stockholm Förhandlingar, 76(4): 517-604. https://doi.org/10.1080/11035895409453581 |
| Lyons, P., Percival, I. G., 2002. Middle to Late Ordovician Age for the Jindalee Group of the Lachlan Fold Belt, New South Wales: Conodont Evidence and some Tectonic Implications. Australian Journal of Earth Sciences, 49(5): 801-808. https://doi.org/10.1046/j.1440-0952.2002.00951.x |
| McTavish, R. A., 1973. Prioniodontacean Conodonts from the Emanuel Formation (Lower Ordovician) of Western Australia. Geologica et Palaeontologica, 7: 27-58 http://www.researchgate.net/publication/281306728_Prioniodontacean_conodonts_from_the_Emanuel_Formation_Lower_Ordovician_of_Western_Australia |
| Murray, S. I., Stewart, I. R., 2001. Palaeogeographic Significance of Ordovician Conodonts from the Lachlan Fold Belt, Southeastern Australia. Historical Biology, 15(1/2): 145-170. https://doi.org/10.1080/10292380109380588 doi: 10.1080/10292380109380588?cookieSet=1 |
| Nielsen, A. T., 2004. Ordovician Sea Level Changes: A Baltoscandian Perspective. In: Webby, B. D., Paris, F., Droser, M. L., et al., eds., The Great Ordovician Biodiversification Event. Columbia University Press. 84-94 |
| Nicoll, R. S., 1980. Middle Ordovician Conodonts from the Pittman Formation, Canberra, A. C. T. BMR Journal of Australian Geology and Geophysics, 5: 150-153 http://ga.gov.au/metadata-gateway/metadata/record/81036/ |
| Och, D. J., Percival, I. G., Leitch, E. C., 2007. Ordovician Conodonts from the Watonga Formation, Port Macquarie, Northeast New South Wales. Proceedings-Linnean Society of New South Wales, 128: 209-216 http://www.researchgate.net/publication/257927766_Ordovician_Conodonts_from_the_Watonga_Formation_Port_Macquarie_Northeast_New_South_Wales |
| Percival, I. G., 2006. Conodont Biostratigraphy and Biogeography of Ordovician Chert from New South Wales, Australia. Programme and Abstracts: International Conodont Symposium 2006, Leicester. 64 |
| Percival, I. G., 2012. Biotic Characteristics of Ordovician Deep-Water Cherts from Eastern Australia. Palaeogeography, Palaeoclimatology, Palaeoecology, 367/368: 63-72. https://doi.org/10.1016/j.palaeo.2011.11.012 http://www.sciencedirect.com/science/article/pii/S0031018211005657 |
| Percival, I. G., Glen, R. A., 2007. Ordovician to Earliest Silurian History of the Macquarie Arc, Lachlan Orogen, New South Wales. Australian Journal of Earth Sciences, 54(2/3): 143-165. https://doi.org/10.1080/08120090601146789 doi: 10.1080/08120090601146789?scroll=top&needAccess=true |
| Percival, I. G., Quinn, C. D., 2011. Reassessment of Lower Palaeozoic Geology West of the Catombal Range, Wellington Region, Central New South Wales. Proceedings-Linnean Society of New South Wales, 132: 221-235 http://www.researchgate.net/publication/257927838_Reassessment_of_Lower_Palaeozoic_geology_west_of_the_Catombal_Range_Wellington_region_central_New_South_Wales |
| Percival, I. G., Zhen, Y. Y., 2007. Darriwilian Conodonts of Eastern Australia: Biostratigraphy and Biogeographic Distribution. Proceedings of the Tenth International Symposium on the Ordovician System, The Third International Symposium on the Silurian System, and IGCP 503 Annual Meeting. June, 2007, Nanjing. Acta Palaeontologica Sinica, 46(Suppl. ): 387-392 http://www.researchgate.net/publication/313218989_Darriwilian_conodonts_of_eastern_Australia_Biostratigraphy_and_biogeographic_distribution |
| Percival, I. G., Kraft, P., Zhang, Y. D., et al., 2015. A Long-Overdue Systematic Revision of Ordovician Graptolite Faunas from New South Wales, Australia. The Ordovician Exposed: Short Papers and Abstracts for 12th International Symposium on the Ordovician System, James Madison University, June 2015, Harrisonburg, Virginia, USA. Stratigraphy, 12(2): 47-53 http://www.researchgate.net/publication/280082982_Stratigraphy_12_2_A_long-overdue_systematic_revision_of_Ordovician_graptolite_faunas_from_New_South_Wales_Australia/download |
| Percival, I. G., Quinn, C. D., Glen, R. A., 2011. Review of Cambrian and Ordovician Stratigraphy in New South Wales. Quarterly Notes of the Geological Survey of New South Wales, 137: 1-39 http://www.researchgate.net/publication/256000673_Review_of_Cambrian_and_Ordovician_stratigraphy_in_New_South_Wales/download |
| Percival, I. G., Zhen, Y. Y., Webby, B. D., 2003. Early Ordovician Conodont Distribution from Craton to Basin and Island Terranes in East Gondwana. In: Albanesi, G. L., Beresi, M. S., Peralta, S. H., eds., Ordovician from the Andes. INSUGEO, Serie Correlación Geológica 17, Tucumán, Argentina. 533-537 |
| Pohler, S. M. L., 1994. Conodont Biofacies of Lower to Lower Middle Ordovician Mega-Conglomerates, Cow Head Group, Western Newfoundland. Geological Survey of Canada, Bulletin, 459: 1-71. https://doi.org/10.4095/194129 http://ci.nii.ac.jp/naid/10021382777 |
| Quinn, C. D., Percival, I. G., Glen, R. A., et al., 2014. Ordovician Marginal Basin Evolution near the Palaeo-Pacific East Gondwana Margin, Australia. Journal of the Geological Society, 171(5): 723-736. https://doi.org/10.1144/jgs2012-034 |
| Rasmussen, J. A., Stouge, S., 2018. Baltoscandian Conodont Biofacies Fluctuations and Their Link to Middle Ordovician (Darriwilian) Global Cooling. Palaeontology, 61(3): 391-416. https://doi.org/10.1111/pala.12348 |
| Repetski, J. E., 1982. Conodonts from E1 Paso Group (Lower Ordovician) of Westernmost Texas and Southern New Mexico. New Mexico Bureau of Mines and Mineral Resources, Memoir, 40: 1-120 |
| Sergeeva, S. P., 1963. Conodonts from the Lower Ordovician of the Leningrad region. Paleontologicheskiy Zhurnal, Akademiya Nauk SSSR, 2: 93-108 http://ci.nii.ac.jp/naid/10021382781 |
| Serpagli, E., 1974. Lower Ordovician Conodonts from Precordilleran Argentina (Province of San Juan). Bollettino Della Societa Paleontologica Italiana, 13(1): 17-98 http://ci.nii.ac.jp/naid/10021382782 |
| Stewart, I. R., 1988. Conodonts. In: Douglas, J. G., Ferguson, J. A., eds., Geology of Victoria. Geological Society of Australia (Victorian Division), Melbourne. 663: 79-81 |
| Stewart, I. R., Fergusson, C. L., 1988. A Lower to Middle Ordovician Age for the Hotham Group, Eastern Victoria. Proceedings of the Royal Society of Victoria, 100: 15-20 http://www.researchgate.net/publication/292655389_A_Lower_to_Middle_Ordovician_age_for_the_Hotham_Group_eastern_Victoria |
| Stewart, I. R., Fergusson, C. L., 1995. Ordovician Conodonts from the Lue Beds, Mudgee and Sunlight Creek Formation, Goulburn, New South Wales. Memoirs of the Association of Australasian Palaeontologists, 18: 164 |
| Stewart, I. R., Glen, R. A., 1986. An Ordovician Age for Part of the Girilambone Group at Yanda Creek, East of Cobar. Geological Survey of New South Wales, Quarterly Notes, 64: 23-25 |
| Stewart, I. R., Glen, R. A., 1991. New Cambrian and Early Ordovician Ages from the New South Wales South Coast. Geological Survey of New South Wales, Quarterly Notes, 85: 1-8 http://www.researchgate.net/publication/292287978_New_Cambrian_and_Early_Ordovician_ages_from_the_New_South_Wales_south_coast |
| Stewart, I. R., Nicoll, R. S., 2004. Multielement Apparatus Structure of the Early Ordovician Conodont Oepikodus evae Lindström from Australia and Sweden. Courier Forschungsinstitut Senckenberg, 245: 361-387 http://www.researchgate.net/publication/285736036_Multielement_apparatus_structure_of_the_Early_Ordovician_conodont_Oepikodus_evae_Lindstrom_from_Australia_and_Sweden |
| Stouge, S., 1984. Conodonts of the Middle Ordovician Table Head Formation, Western Newfoundland. Fossils and Strata, 16: 1-145 http://www.sciencedirect.com/science/article/pii/0012825286900073 |
| Stouge, S., 2012. Middle Ordovician (Late Dapingian-Darriwilian) Conodonts from the Cow Head Group and Lower Head Formation, Western Newfoundland, Canada. Canadian Journal of Earth Sciences, 49(1): 59-90. https://doi.org/10.1139/e11-057 |
| Stouge, S., Bagnoli, G., 1988. Early Ordovician Conodonts from Cow Head Peninsula, Western Newfoundland. Palaeontographica Italica, 75: 89-179 http://ci.nii.ac.jp/naid/10021382788 |
| Stouge, S., Bagnoli, G., McIlroy, D., 2017. Cambrian-Middle Ordovician Platform-Slope Stratigraphy, Palaeontology and Geochemistry of Western Newfoundland, Canada. International Symposium on the Ediacaran-Cambrian Transition. June 2017, St. John's, Newfoundland, Field Trip 2. Open File NFLD/3325. 106 |
| Sweet, W. C., Bergström, S. M., 1962. Conodonts from the Pratt Ferry Formation (Middle Ordovician) of Alabama. Journal of Paleontology, 36: 1214-1252 http://www.onacademic.com/detail/journal_1000038708161510_4974.html |
| Tolmacheva, T. Y., 2014. Biostratigraphy and Biogeography of Ordovician Conodonts from the Western Part of the Central Asian Fold Belt. Trudy VSEGEI, Novaya Seriya, 356: 1-263 (in Russian) http://www.researchgate.net/publication/311739137_Biostratigraphy_nd_biogeography_of_the_Ordovician_conodonts_from_the_western_part_of_Central_Asian_Fold_Belt_Biostratigrafia_i_biogeografia_konodontov_ordovika_zapadnoj_casti_Centralno-Aziatskogo_Skla |
| Tolmacheva, T. Y., 2019. Biostratigraphy and Biodiversity of the Ordovician Conodonts from the Deep-Water Siliceous Deposits of Western Part of Central-Asian Fold Belt. In: Obut, O. T., Sennikov, N. V., Kipriyanova, T. P., eds., 13th International Symposium on the Ordovician System: Contributions of International Symposium. July 19-22, 2019, Novosibirsk. 203-205 |
| Tolmacheva, T., Löfgren, A., 2000. Morphology and Paleogeography of the Ordovician Conodont Paracordylodus gracilis Lindström, 1955: Comparison of Two Populations. Journal of Paleontology, 74(6): 1114-1121 doi: 10.1017/S0022336000017650 |
| Tolmacheva, T. Y., Purnell, M. A., 2002. Apparatus Composition, Growth, and Survivorship of the Lower Ordovician Conodont Paracordylodus gracilis Lindstrom, 1955. Palaeontology, 45(2): 209-228. https://doi.org/10.1111/1475-4983.00234 |
| Tolmacheva, T., Holmer, L., Popov, L., et al., 2004. Conodont Biostratigraphy and Faunal Assemblages in Radiolarian Ribbon-Banded Cherts of the Burubaital Formation, West Balkhash Region, Kazakhstan. Geological Magazine, 141(6): 699-715. https://doi.org/10.1017/s0016756804009902 doi: 10.1017/S0016756804009902 |
| Tolmacheva, T. Y., Degtyarev, K. E., Samuelsson, J., et al., 2008. Middle Cambrian to Lower Ordovician Faunas from the Chingiz Mountain Range, Central Kazakhstan. Alcheringa: An Australasian Journal of Palaeontology, 32(4): 443-463. https://doi.org/10.1080/03115510802418099 |
| Tolmacheva, T. Y., Degtyarev, K. E., Ryazantsev, A. V., et al., 2009. Conodonts from the Upper Ordovician Siliceous Rocks of Central Kazakhstan. Paleontological Journal, 43(11): 1498-1512. https://doi.org/10.1134/s0031030109110136 doi: 10.1134/S0031030109110136 |
| Trotter, J. A., Webby, B. D., 1994. Upper Ordovician Conodonts from the Malongulli Formation, Cliefden Caves Area, Central New South Wales. AGSO Journal of Australian Geology and Geophysics, 15(4): 475-499 http://www.researchgate.net/publication/293208927_Upper_Ordovician_conodonts_from_the_Malongulli_Formation_Cliefden_Caves_area_central_New_South_Wales |
| Wang, Z. H., Bergström, S. M., Zhen, Y. Y., et al., 2013. Ordovician Conodonts from Dashimen, Wuhai in Inner Mongolia and the Significance of the Discovery of the Histiodella Fauna. Acta Micropalaeontologica Sinica, 30(4): 323-343 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-WSGT201304001.htm |
| Wang, Z. H., Zhen, Y. Y., Bergström, S. M., et al., 2019. A New Conodont Biozone Classification of the Ordovician System in South China. Palaeoworld, 28(1/2): 173-186. https://doi.org/10.1016/j.palwor.2018.09.002 http://www.sciencedirect.com/science/article/pii/S1871174X18300131 |
| Watson, S. T., 1988. Ordovician Conodonts from the Canning Basin (W. Australia). Palaeontographica Abteilung A, 203(4-6): 91-147 http://www.researchgate.net/publication/288296905_Ordovician_conodonts_from_the_Canning_Basin_W_Australia |
| Wu, R. C., Stouge, S., Percival, I. G., et al., 2014. Early-Middle Ordovician Conodont Biofacies on the Yangtze Platform Margin, South China: Applications to Palaeoenvironment and Sea-Level Changes. Journal of Asian Earth Sciences, 96: 194-204. https://doi.org/10.1016/j.jseaes.2014.09.003 |
| VandenBerg, A. H. M., Cooper, R. A., 1992. The Ordovician Graptolite Sequence of Australasia. Alcheringa: An Australasian Journal of Palaeontology, 16(1): 33-85. https://doi.org/10.1080/03115519208619032 |
| VandenBerg, A. H. M., Stewart, I. R., 1992. Ordovician Terranes of the Southeastern Lachlan Fold Belt: Stratigraphy, Structure and Palaeogeographic Reconstruction. Tectonophysics, 214(1/2/3/4): 159-176. https://doi.org/10.1016/0040-1951(92)90195-c http://www.sciencedirect.com/science/article/pii/004019519290195C |
| Zhang, Y. D., Zhan, R. B., Zhen, Y. Y., et al., 2019. Ordovician Integrative Stratigraphy and Timescale of China. Science China Earth Sciences, 62(1): 61-88. https://doi.org/10.1007/s11430-017-9279-0 |
| Zhen, Y. Y., 2020. Revision of the Darriwilian (Middle Ordovician) Conodonts Documented by Watson (1988) from Subsurface Canning Basin, Western Australia. Alcheringa: An Australasian Journal of Palaeontology, 44(2): 217-252. https://doi.org/10.1080/03115518.2020.1737227 |
| Zhen, Y. Y., Percival, I. G., 2003. Ordovician Conodont Biogeography-Reconsidered. Lethaia, 36(4): 357-369 doi: 10.1080/00241160310006402 |
| Zhen, Y. Y., Percival, I. G., 2004a. Darriwilian (Middle Ordovician) Conodonts from the Weemalla Formation, South of Orange, New South Wales. Memoirs of the Association of Australasian Palaeontologists, 30: 153-178 http://www.researchgate.net/publication/301559124_Darriwilian_Middle_Ordovician_conodonts_from_the_Weemalla_Formation_south_of_Orange_New_South_Wales |
| Zhen, Y. Y., Percival, I. G., 2004b. Middle Ordovician (Darriwilian) Conodonts from Allochthonous Limestones in the Oakdale Formation of Central New South Wales. Alcheringa: An Australasian Journal of Palaeontology, 28(1): 77-110. https://doi.org/10.1080/03115510408619276 |
| Zhen, Y. Y., Percival, I. G., 2017. Late Ordovician Conodont Biozonation of Australia-Current Status and Regional Biostratigraphic Correlations. Alcheringa: An Australasian Journal of Palaeontology, 41(3): 285-305. https://doi.org/10.1080/03115518.2017.1282982 |
| Zhen, Y. Y., Pickett, J. W., 2008. Ordovician (Early Darriwilian) Conodonts and Sponges from West of Parkes, Central New South Wales. Proceedings of the Linnean Society of New South Wales, 129: 57-82 http://www.researchgate.net/publication/292698551_Ordovician_early_Darriwilian_conodonts_and_sponges_from_west_of_Parkes_central_New_South_Wales |
| Zhen, Y. Y., Rutledge, J., 2020. Conodont Biostratigraphy of the Ordovician Turbiditic Successions in New South Wales. Geological Survey of New South Wales, Report. No. GS2020/0459 |
| Zhen, Y. Y., Cooper, R. A., Simes, J. E., et al., 2011. Darriwilian (Middle Ordovician) Conodonts from the Maruia-Springs Junction Area, New Zealand. Memoirs of the Association of Australasian Palaeontologists, 42(42): 285-319 http://www.researchgate.net/publication/257927916_Darriwilian_Middle_Ordovician_conodonts_from_the_Maruia-_Springs_Junction_area_New_Zealand |
| Zhen, Y. Y., Laurie, J. R., Percival, I. G., et al., in Press. Ordovician Conodonts from the Horn Valley Siltstone of the Amadeus Basin, Central Australia. Australasian Palaeontological Memoirs, 56 |
| Zhen, Y. Y., Nicoll, R. S., Percival, I. G., et al., 2001. Ordovician Rhipidognathid Conodonts from Australia and Iran. Journal of Paleontology, 75(1): 186-207. https://doi.org/10.1017/s002233600003198x doi: 10.1666/0022-3360(2001)075<0186:ORCFAA>2.0.CO;2 |
| Zhen, Y. Y., Percival, I. G., Webby, B. D., 2004. Early Ordovician (Bendigonian) Conodonts from Central New South Wales, Australia. Courier Forschungsinstitut Senckenberg, 245: 39-73 http://ci.nii.ac.jp/naid/10021382799 |
| Zhen, Y. Y., Percival, I. G., Liu, J. B., 2006. Early Ordovician Triangulodus (Conodonta) from the Honghuayuan Formation of Guizhou, South China. Alcheringa: An Australasian Journal of Palaeontology, 30(2): 191-212. https://doi.org/10.1080/03115510608619313 |
| Zhen, Y. Y., Percival, I. G., Liu, J. B., et al., 2009. Conodont Fauna and Biostratigraphy of the Honghuayuan Formation (Early Ordovician) of Guizhou, South China. Alcheringa: An Australasian Journal of Palaeontology, 33(3): 257-295. https://doi.org/10.1080/03115510903043655 |
| Zhen, Y. Y., Webby, B. D., Barnes, C. R., 1999. Upper Ordovician Conodonts from the Bowan Park Succession, Central New South Wales, Australia. Geobios, 32(1): 73-104 doi: 10.1016/S0016-6995(99)80084-9 |
| Zhen, Y. Y., Zhang, Y. D., Tang, Z. C., et al., 2015. Early Ordovician Conodonts from Zhejiang Province, Southeast China and Their Biostratigraphic and Palaeobiogeographic Implications. Alcheringa: An Australasian Journal of Palaeontology, 39(1): 109-141. https://doi.org/10.1080/03115518.2015.958295 |
| Zhen, Y. Y., Zhang, Y. D., Percival, I. G., et al., 2020. Basal Darriwilian Graptolites and Associated Conodonts from New South Wales and Their Biostratigraphic Implications. Quarterly Notes of the Geological Survey of New South Wales, 153: 1-17 http://www.researchgate.net/publication/344663941_Basal_Darriwilian_graptolites_and_associated_conodonts_from_New_South_Wales_and_their_biostratigraphic_implications |
Figures(7)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: