| Citation: | Hanxiao Li, Miaoyan Wang, Muhui Zhang, Paul B. Wignall, Manuel Rigo, Yanlong Chen, Xianlang Wu, Zhumin Ouyang, Baojin Wu, Zhaoyang Yi, Zaitian Zhang, Xulong Lai. First Records of Late Triassic Conodont Fauna and δ13Ccarb from the Dengdengqiao Section, Dangchang County, Gansu Province, Northwestern China. Journal of Earth Science, 2021, 32(3): 646-656. doi: 10.1007/s12583-021-1428-9

|

| Balini, M., Lucas, S. G., Jenks, J. F., et al., 2010. Triassic Ammonoid Biostratigraphy: An Overview. Geological Society, London, Special Publications, 334(1): 221-262. https://doi.org/10.1144/sp334.10 |
| Bernhard, J. M., 1986. Characteristic Assemblages and Morphologies of Benthic Foraminifera from Anoxic Organic-Rich Deposits: Jurassic through Holocene. Journal of Foraminiferal Research, 16(3): 207-215. https://doi.org/10.2113/gsjfr.16.3.207 |
| Channell, J. E. T., Kozur, H. W., Sieversa, T., et al., 2003. Carnian-Norian Biomagnetostratigraphy at Silická Brezová (Slovakia): Correlation to Other Tethyan Sections and to the Newark Basin. Palaeogeography, Palaeoclimatology, Palaeoecology, 191(2): 65-109. https://doi.org/10.1016/S0031-0182(02)006545 |
| Chen, Y. L., Krystyn, L., Orchard, J. M., et al., 2016. A Review of the Evolution, Biostratigraphy, Provincialism and Diversity of Middle and Early Late Triassic Conodonts. Papers in Palaeontology, 2(2): 235-263. https://doi.org/10.1002/spp2.1038 |
| Chen, Y. L., Lukeneder, A., 2017. Late Triassic (Julian) Conodont Biostratigraphy of a Transition from Reefal Limestone to Deep-Water Environments on the Cimmerian Terranes (Taurus Mountains, Southern Turkey). Papers in Palaeontology, 3(3): 441-460. https://doi.org/10.1002/spp2.1082 |
| Chen, Y., Ye, Q., Jiang, H. S., et al., 2019. Conodonts and Carbon Isotopes during the Permian-Triassic Transition on the Napo Platform, South China. Journal of Earth Science, 30(2): 244-257. https://doi.org/10.1007/s12583-018-0884-3 |
| Chen, Y. L., Scholze, F., Richoz, S., et al., 2019. Middle Triassic Conodont Assemblages from the Germanic Basin: Implications for Multi-Element Taxonomy and Biogeography. Journal of Systematic Palaeontology, 17(5): 359-377. https://doi.org/10.1080/14772019.2018.1424260 |
| Dong, Y. P., Zhang, X. N., Liu, X. M., et al., 2015. Propagation Tectonics and Multiple Accretionary Processes of the Qinling Orogen. Journal of Asian Earth Sciences, 104: 84-98. https://doi.org/10.1016/j.jseaes.2014.10.007 |
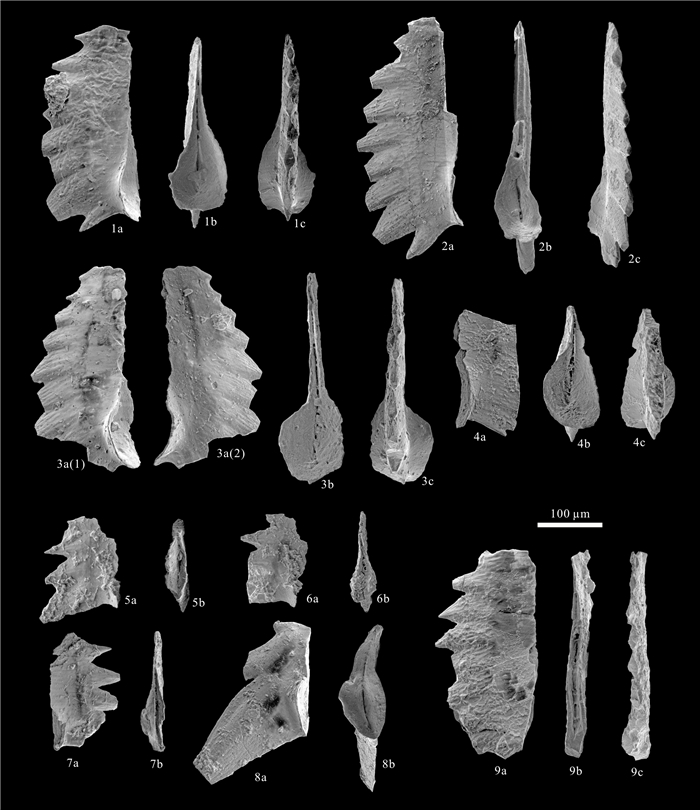
| Fåhraeus, L. E., Ryley, C. C., 1989. Multielement Species of Misikella Kozur and Mock, 1974 and Axiothea n. gen. (Conodonta) from the Mamonia Complex (Upper Triassic), Cyprus. Canadian Journal of Earth Sciences, 26(6): 1255-1263. https://doi.org/10.1139/e89-106 |
| Gallet, Y., Besse, J., Krystyn, L., et al., 1994. Magnetostratigraphy of the Mayerling Section (Austria) and Erenkolu Mezarlik (Turkey) Section: Improvement of the Carnian (Late Triassic) Magnetic Polarity Time Scale. Earth and Planetary Science Letters, 125(1-4): 173-191. https://doi.org/10.1016/0012-821X(94)90214-3 |
| Jiang, H. S., Yuan, J. L., Chen, Y., et al., 2019. Synchronous Onset of the Mid-Carnian Pluvial Episode in the East and West Tethys: Conodont Evidence from Hanwang, Sichuan, South China. Palaeogeography, Palaeoclimatology, Palaeoecology, 520: 173-180. https://doi.org/10.1016/j.palaeo.2019.02.004 |
| Ishida, K., Hirsch, F., 2011. The Triassic Conodonts of the NW Malayan Kodiang Limestone Revisited: Taxonomy and Paleogeographic Significance. Gondwana Research, 19: 22-36. https://doi.org/10.1016/j.gr.2010.05.008 |
| Karádi, V., Kozur, H., Görög, Á., 2013. Stratigraphically Important Lower Norian Conodonts from the Csővár Borehole (Csv-1), Hungary-Comparison with the Conodont Succession of the Norian GSSP Candidate Pizzo Mondello (Sicily, Italy). New Mexico Museum of Natural History and Science Bulletin, 61: 284-295 http://www.researchgate.net/publication/256115961_Stratigraphically_important_lower_Norian_conodonts_from_the_Csovar_borehole_Csv-1_Hungary_-_comparison_with_the_conodont_succession_of_the_Norian_GSSP_candidate_PizzoMondello_Sicily_Italy |
| Karádi, V., Cau, A., Mazza, M., et al., 2020. The Last Phase of Conodont Evolution during the Late Triassic: Integrating Biostratigraphic and Phylogenetic Approaches. Palaeogeography, Palaeoclimatology, Palaeoecology, 549: 109144. https://doi.org/10.1016/j.palaeo.2019.03.045 |
| Kolar-Jurkovšek, T., Jurkovšek, B., 2010. New Paleontological Evidence of the Carnian Strata in the Mežica Area (Karavanke Mts, Slovenia): Conodont Data for the Carnian Pluvial Event. Palaeogeography, Palaeoclimatology, Palaeoecology, 290(1-4): 81-88. https://doi.org/10.1016/j.palaeo.2009.06.015 |
| Kolar-Jurkovšek, T., Jurkovšek, B., 2019. Konodonti Slovenije (Conodonts of Slovenia). Geološki zavod Slovenije, Ljubljana. 260 |
| Kozur, H., 1993. Nicoraella postkockeli n. sp., a New Conodont Species from the Lower Carnian of Hungary. Neues Jahrbuch für Geologie und Paläontologie Monatshefte, 7: 405-412 |
| Kozur, H., 2003. Integrated Ammonoid, Conodont and Radiolarian Zonation of the Triassic. Hallesches Jahrbuch Geowissenschaften, B25: 49-79 |
| Kozur, H., Mock, R., 1991. New Middle Carnian and Rhaetian Conodonts from Hungary and the Alps. Stratigraphic Importance and Tectonic Implications for the Buda Mountains and Adjacent Areas. Jahrbuch der Geologischen Bundesanstalt, 134(2): 271-297 http://ci.nii.ac.jp/naid/10003918868 |
| Kozur, H., Mock, R., 1974. Zwei neue Conodonten-Arten aus der Trias des Slowakischen Karstes. Časopis pro Mineralogii a Geologii, 19(2): 135-139 (in German) http://www.researchgate.net/publication/288028155_Zwei_neue_Conodonten-Arten_aus_der_Trias_des_Slowakischen_Karstes |
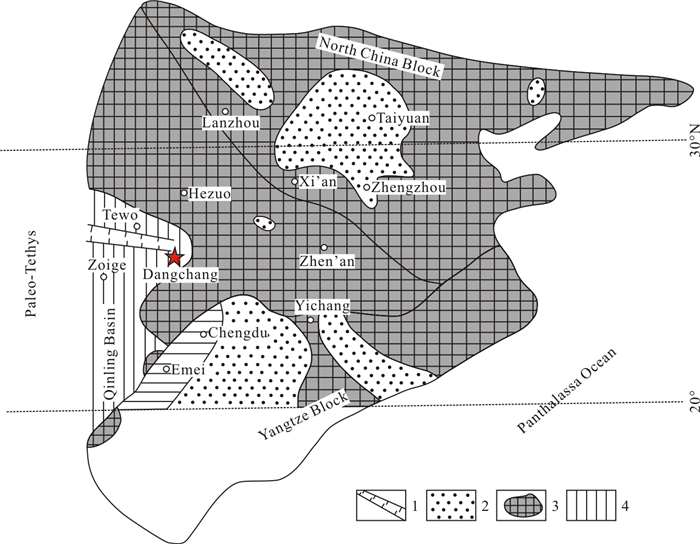
| Lai, X. L., 1992. Qinyu-Dengdengqiao, Daheba-Xinchengzi Section in Dangchang County, Gansu Province. In: Yin, H. F., Yang, F. Q., Huang, Q. S., et al., eds., Triassic in Qinling and Adjacent Areas. China University of Geosciences Press, Wuhan. 20-29 (in Chinese) |
| Lai, X. L., Yin, H. F., Yang, F. Q., 1992. Reconstruction of the Triassic Qinling Sea. Exploration of Geosciences, (6): 57-64 |
| Lai, X. L., Yin, H. F., Yang, F. Q., 1995. Reconstruction of the Qinling Triassic Paleo-Ocean. Earth Science, 20(6): 648-656 (in Chinese with English Abstract) http://www.researchgate.net/publication/271907717_Reconstruction_of_Qinling_Triassic_Paleo-ocean |
| Lehrmann, D. J., Stepchinski, L., Altiner, D., et al., 2015. An Integrated Biostratigraphy (Conodonts and Foraminifers) and Chronostratigraphy (Paleomagnetic Reversals, Magnetic Susceptibility, Elemental Chemistry, Carbon Isotopes and Geochronology) for the Permian-Upper Triassic Strata of Guandao Section, Nanpanjiang Basin, South China. Journal of Asian Earth Sciences, 108: 117-135. https://doi.org/10.1016/j.jseaes.2015.04.030 |
| Li, H. X., Jiang, H. S., Chen, Y. L., et al., 2019. Smithian Platform-Bearing Gondolellid Conodonts from Yiwagou Section, Northwestern China and Implications for Their Geographic Distribution in the Early Triassic. Journal of Paleontology, 93(3): 496-511. https://doi.org/10.1017/jpa.2018.93 |
| Li, Y. J., Zhao, R. F., Liu, Z. W., et al., 2003. Triassic Sedimentation and Basin Evolution in the Western Qinling. Geology in China, 30(3): 268-273 (in Chinese with English Abstract) http://d.wanfangdata.com.cn/Periodical/zgdizhi200303007 |
| Liu, S., Sun, Z. Y., Ji, C., et al., 2020. Conodont Biostratigraphy and Age of the Early Triassic Fish-Bearing-Nodule Levels from Nanjing and Jurong, Jiangsu Province, South China. Journal of Earth Science, 31(1): 9-22. https://doi.org/10.1007/s12583-019-1232-y |
| Lyu, Z. Y., Orchard, M. J., Chen, Z. Q., et al., 2018. A Taxonomic Re-Assessment of the Novispathodus waageni Group and Its Role in Defining the Base of the Olenekian (Lower Triassic). Journal of Earth Science, 29(4): 824-836. https://doi.org/10.1007/s12583-018-0795-3 |
| Mazza, M., Cau, A., Rigo, M., 2012a. Application of Numerical Cladistic Analyses to the Carnian-Norian Conodonts: A New Approach for Phylogenetic Interpretations. Journal of Systematic Palaeontology, 10(3): 401-422. https://doi.org/10.1080/14772019.2011.573584 |
| Mazza, M., Rigo, M., Gullo, M., 2012b. Taxonomy and Biostratigraphic Record of the Upper Triassic Conodonts of the Pizzo Mondello Section (Western Sicily, Italy), GSSP Candidate for the Base of the Norian. Rivista Italiana di Paleontologia e Stratigrafia, 118(1): 85-130 |
| Mosher, L. C., 1968. Triassic Conodonts from Western North America and Europe and Their Correlation. Journal of Paleontology, 42: 895-946 |
| O'Dogherty, L., Carter, E. S., Gorican, S., et al., 2010. Triassic Radiolarian Biostratigraphy. In: Lucas, S. G., ed., The Triassic Timescale. Special Publication of the Geological Society of London, 334: 163-200. https://doi.org/10.1144/sp334.8 |
| Orchard, M. J., 1991. Late Triassic Conodont Biochronology and Biostratigraphy of the Kunga Group, Queen Charlotte Islands, British Columbia. In: Woodsworth, G. W., ed., Evolution and Hydrocarbon Potential of the Queen Charlotte Basin, British Columbia. Geological Survey of Canada Paper, 90-10: 173-193 |
| Orchard, M. J., 2010. Triassic Conodonts and Their Role in Stage Boundary Definition. Geological Society, London, Special Publications, 334(1): 139-161. https://doi.org/10.1144/sp334.7 |
| Orchard, M. J., 2014. Conodonts from the Carnian-Norian Boundary (Upper Triassic) of Black Bear Ridge, Northeastern British Columbia, Canada. New Mexico Museum of Natural History and Science Bulletin, 64: 1-139 http://www.researchgate.net/publication/292234849_Conodonts_from_the_Carnian-Norian_boundary_Upper_Triassic_of_Black_Bear_Ridge_Northeasern_British_Columbia_Canada |
| Orchard, M. J., Balini, M., 2007. Conodonts from the Ladinian-Carnian Boundary Beds of South Canyon, New Pass Range, Nevada, USA. New Mexico Museum of Natural History and Science Bulletin, 41: 333-340 http://www.researchgate.net/publication/260640392_Conodonts_from_the_Ladinian-Carnian_boundary_beds_of_South_Canyon_New_Pass_Range_Nevada_USA |
| Orchard, M. J., Tozer, E. T., 1997. Triassic Conodont Biochronology, Its Calibration with the Ammonoid Standard, and a Biostratigraphic Summary for the Western Canada Sedimentary Basin. Bulletin of Canadian Petroleum Geology, 45(4): 675-692. https://doi.org/10.35767/gscpgbull.45.4.675 |
| Rigo, M., Preto, N., Roghi, G., et al., 2007. A Rise in the Carbonate Compensation Depth of Western Tethys in the Carnian (Late Triassic): Deep-Water Evidence for the Carnian Pluvial Event. Palaeogeography, Palaeoclimatology, Palaeoecology, 246(2-4): 188-205. https://doi.org/10.1016/j.palaeo.2006.09.013 |
| Rigo, M., Mazza, M., Karádi, V., et al., 2018. New Upper Triassic Conodont Biozonation of the Tethyan Realm. In: Tanner, L. H., ed., The Late Triassic World. Topics in Geobiology, 46: 189-235. https://doi.org/10.1007/978-3-319-68009-5_6 |
| Ryley, C. C., 1987. Multielement Taxonomy, Biostratigraphy, and Paleoecology of Late Triassic Conodonts from the Mamonia Complex, Southwestern Cyprus: [Dissertation]. Memorial University of Newfoundland, St. Johnʼs, NL, Canada. 1-191 |
| Shi, Z., Preto, N., Jiang, H., et al., 2017. Demise of Late Triassic Sponge Mounds along the Northwestern Margin of the Yangtze Block, South China: Related to the Carnian Pluvial Phase?. Palaeogeography, Palaeoclimatology, Palaeoecology, 474: 247-263. https://doi.org/10.1016/j.palaeo.2016.10.031 |
| Song, H. Y., Tong, J. N., Algeo, T. J., et al., 2013. Large Vertical δ13CDIC Gradients in Early Triassic Seas of the South China Craton: Implications for Oceanographic Changes Related to Siberian Traps Volcanism. Global and Planetary Change, 105: 7-20. https://doi.org/10.1016/j.gloplacha.2012.10.023 |
| Sudar, M., Budurov, K., 1979. New Conodonts from the Triassic in Yugoslavia and Bulgaria. Geologica Balcanica, 9: 47-52 http://www.researchgate.net/publication/284978298_New_conodonts_from_the_Triassic_in_Yugoslavia_and_Bulgaria |
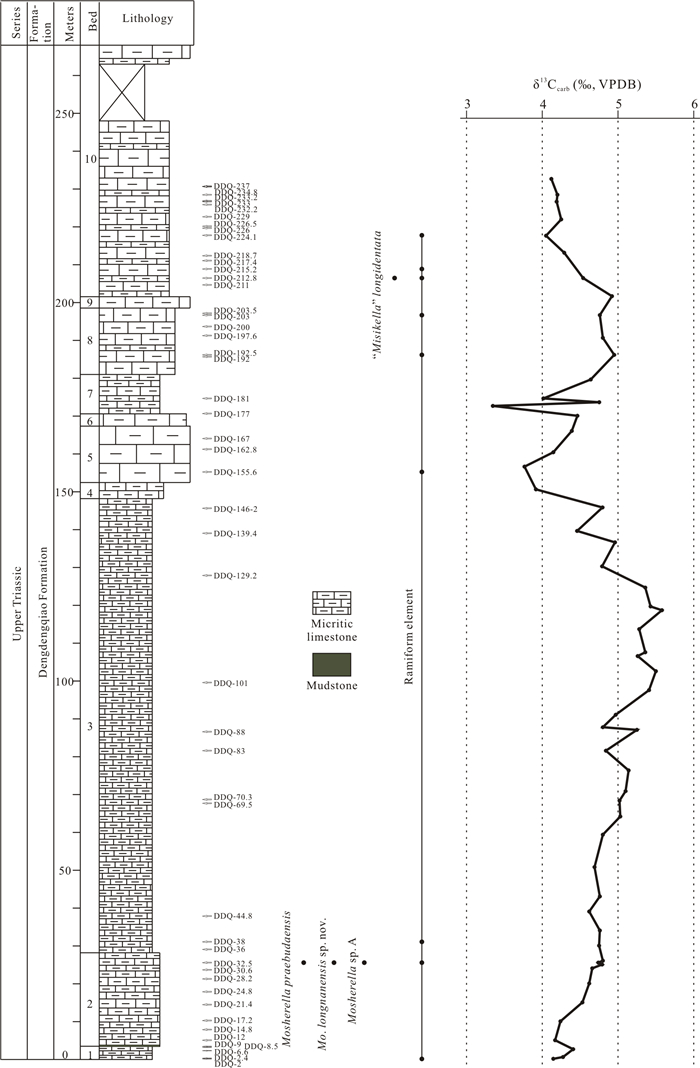
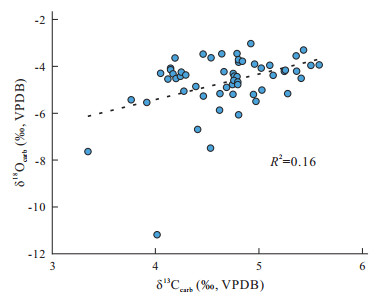
| Sun, Y. D., Wignall, P. B., Joachimski, M. M., et al., 2016. Climate Warming, Euxinia and Carbon Isotope Perturbations during the Carnian (Triassic) Crisis in South China. Earth and Planetary Science Letters, 444: 88-100. https://doi.org/10.1016/j.epsl.2016.03.037 |
| Sweet, W. C., Mosher, L. C., Clark, D. L., et al., 1971. Conodont Biostratigraphy of the Triassic. Geological Society of America Memoirs, 127: 441-466 |
| Tian, C. R., Dai, J. Y., Tian, S. G., 1983. Triassic Conodonts. In: Chengdu Institute of Geology and Mineral Resources, ed., Paleontological Atlas of Southwest China, Volume of Microfossils (Pt. 4, Micropaleontology). Geological Publishing House, Beijing. 345-398 (in Chinese) |
| Wang, H. M., Wang, X. L., Li, R. X., et al., 2005. Triassic Conodont Succession and Stage Subdivision of the Guandao Section, Bianyang, Luodian, Guizhou. Acta Palaeontologica Sinica, 44(4): 611-626 (in Chinese with English Abstract) http://europepmc.org/abstract/cba/599676 |
| Xu, X. R., 1992. Radiolarian. In: Yin, H. F., Yang, F. Q., Huang, Q. S., et al., eds., Triassic in Qinling and Adjacent Areas. China University of Geosciences Press, Wuhan. 68-69 (in Chinese) |
| Yang, B., Li, H. X., Wignall, P. B., et al., 2019. Latest Wuchiapingian to Earliest Triassic Conodont Zones and δ13Ccarb Isotope Excursions from Deep-Water Sections in Western Hubei Province, South China. Journal of Earth Science, 30(5): 1059-1074. https://doi.org/10.1007/s12583-019-1018-2 |
| Yang, Z. H., Guo, J. F., Su, S. R., et al., 2002. New Advances in the Geological Study of the Qinling Orogen. Geology in China, 29(3): 246-256 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-DIZI200203002.htm |
| Yang, Z. H., Chao, H. X., Wu, X., et al., 2016. The Structural Characteristics of Qinling Intra-Continental Orogenic Belt and Its Choula (Drawing-out) Tectonic Orogenic Model. Earth Science Frontiers, 23(4): 63-71 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-DXQY201604006.htm |
| Yin, H. F., Yang, F. Q., Lai, X. L., 1988. Triassic Belts and Indosinian Development of the Qinling Mountains. Geoscience, 2: 355-365 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-XDDZ198803009.htm |
| Yuan, J., Jiang, H., Wang, D., 2015. LST: A New Inorganic Heavy Liquid Used in Conodont Separation. Geological Science and Technology Information, 34(5): 225-230 (in Chinese with English Abstract) http://www.en.cnki.com.cn/Article_en/CJFDTotal-DZKQ201505035.htm |
| Zhang, Z. T., Sun, Y. D., Lai, X. L., 2017. Early Carnian Conodont Fauna at Yongyue, Zhenfeng Area and Its Implication for Ladinian-Carnian Subdivision in Guizhou, South China. Palaeogeography, Palaeoclimatology, Palaeoecology, 486(15): 142-157. https://doi.org/10.1016/j.palaeo.2017.02.011 |
| Zhang, Z. T., Sun, Y. D., Lai, X. L., 2018a. Progresses on Carnian (Late Triassic) Conodont Study in Southwest China. Earth Science, 43(11): 3955-3975 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTotal-DQKX201811013.htm |
| Zhang, Z. T., Sun, Y. D., Lai, X. L., et al., 2018b. Carnian (Late Triassic) Conodont Faunas from South-western China and Their Implications. Papers in Palaeontology, 4(4): 513-535. https://doi.org/10.1002/spp2.1116 |
Figures(7) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: