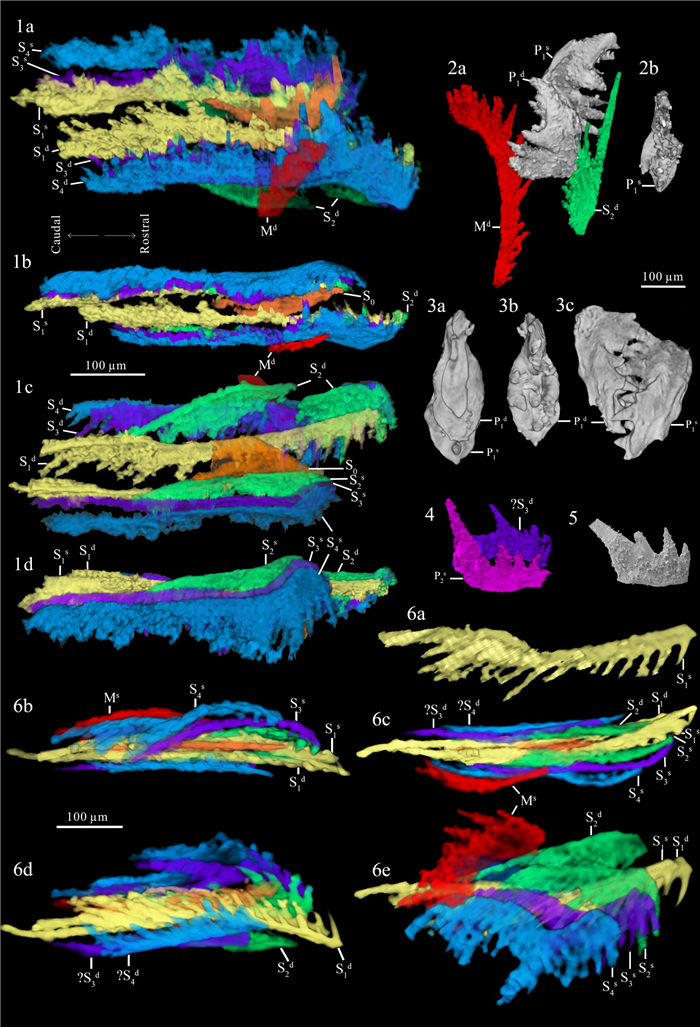
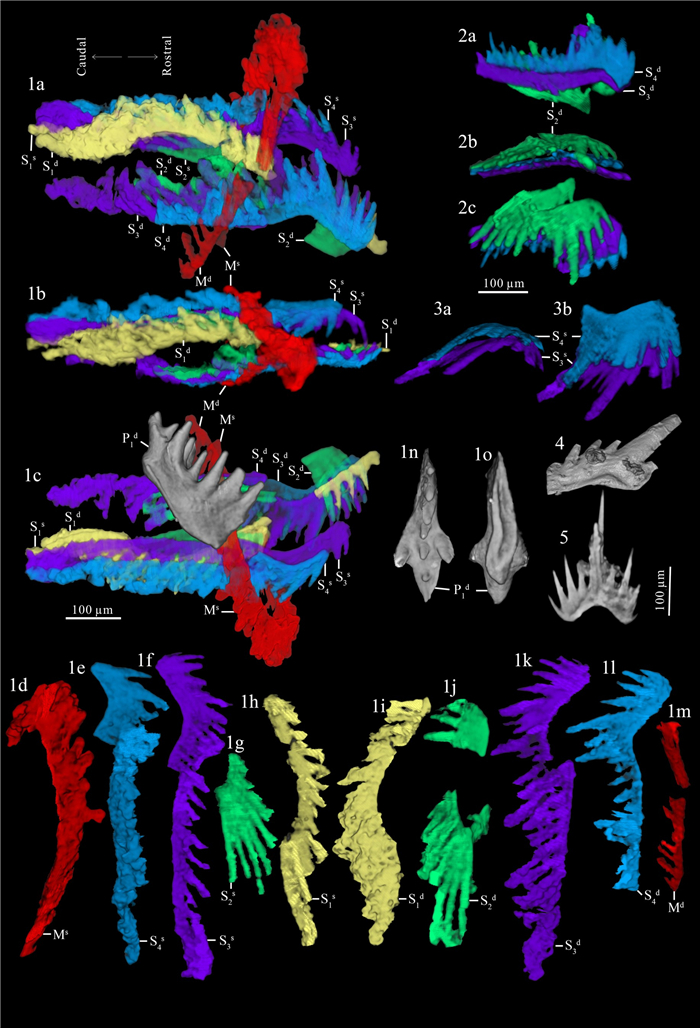
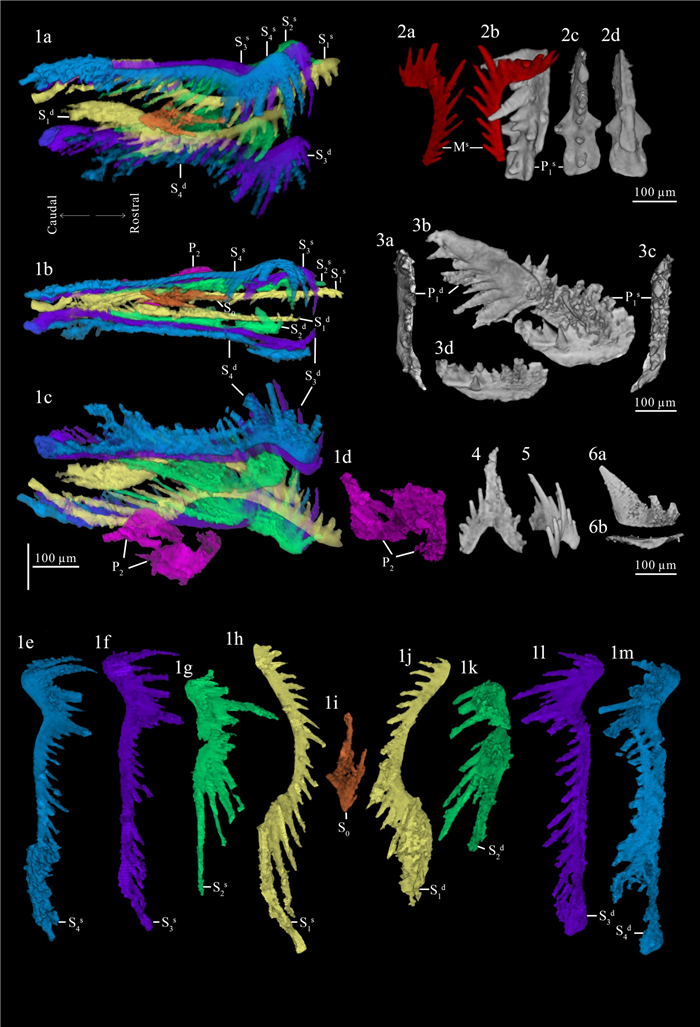
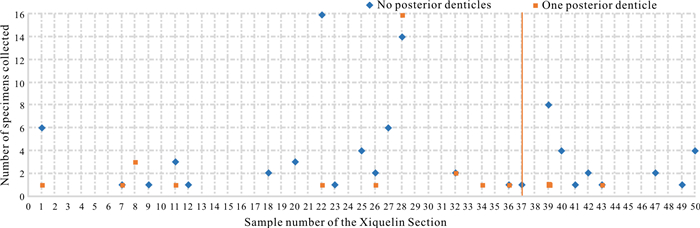
| Citation: | Weiping Zeng, Mark A. Purnell, Haishui Jiang, Muhui Zhang. Late Triassic (Norian) Conodont Apparatuses Revealed by Conodont Clusters from Yunnan Province, Southwestern China. Journal of Earth Science, 2021, 32(3): 709-724. doi: 10.1007/s12583-021-1459-2

|

| Agematsu, S., Sano, H., Sashida, K., et al., 2014. Natural Assemblages of Hindeodus Conodonts from a Permian-Triassic Boundary Sequence, Japan. Palaeontology, 57(6): 1277-1289. https://doi.org/10.1111/pala.12114 |
| Agematsu, S., Uesugi, K., Sano, H., et al., 2017. Reconstruction of the Multielement Apparatus of the Earliest Triassic Conodont, Hindeodus parvus, Using Synchrotron Radiation X-Ray Micro-Tomography. Journal of Paleontology, 91(6): 1220-1227. https://doi.org/10.1017/jpa.2017.61 |
| Aldridge, R. J., Theron, J. N., 1993. Conodonts with Preserved Soft Tissue from a New Ordovician Konservat-Lagerstatte. Journal of Micropalaeontology, 12(1): 113-117. https://doi.org/10.1144/jm.12.1.113 |
| Aldridge, R. J., Purnell, M. A., Gabbott, S. E., et al., 1995. The Apparatus Architecture and Function of Promissum pulchrum Kovács-Endrödy (Conodonta, Upper Ordovician) and the Prioniodontid Plan. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences, 347(1321): 275-291. https://doi.org/10.1098/rstb.1995.0027 |
| Atakul-Özdemir, A., Purnell, M. A., Riley, N. J., 2012. Cladistic Tests of Monophyly and Relationships of Biostratigraphically Significant Conodonts Using Multielement Skeletal Data-Lochriea homopunctatus and the Genus Lochriea. Palaeontology, 55(6): 1279-1291. https://doi.org/10.1111/j.1475-4983.2012.01190.x |
| Bao, J. F., Zhao, Y. J., Wang, C. P., et al., 2012. The Discovery and Significance on the Seismites of Dashuitang Formation in the Upper Triassic Series in Baoshan Jinji Area of Western Yunnan. Acta Sedimentologica Sinica, 30(3): 490-500 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-CJXB201203010.htm |
| BGMRY (Bureau of Geology and Mineral Resources of Yunnan Province), 1980. Geological Map Sheet Baoshan with Explanation, Scale 1: 200 000. Geological Publishing House, Beijing (in Chinese) |
| BGMRY, 1990. Regional Geology of Yunnan Province. Geological Publishing House, Beijing. 728 (in Chinese) |
| BGMRY, 2008. Geological Map Sheet Dali with Explanation, Scale 1: 250 000. Geological Publishing House, Beijing (in Chinese) |
| Briggs, D. E. G., Clarkson, E. N. K., Aldridge, R. J., 1983. The Conodont Animal. Lethaia, 16(1): 1-14. https://doi.org/10.1111/j.1502-3931.1983.tb01993.x |
| Budai, T., Kovács, S., 1986. Contributions to the Stratigraphy of the Rezi Dolomite Formation. M. ÁII Földtani Intézet Jelentése az, 1984: 175-191 http://www.getcited.org/pub/102918168 |
| Budurov, K., Sudar, M. N., 1990. Late Triassic Conodont Stratigraphy. Courier Forschungsinstitut Senckenberg, 118: 203-239 |
| Demo, F., 2017. Analysis of Synchrotron Microtomography 3D Images of Exceptional Fused Cluster of Mockina Slovakensis (Conodonts): [Dissertation]. Università degli Studi di Padova, Padua |
| Donoghue, P. C. J., Purnell, M. A., 1999. Mammal-Like Occlusion in Conodonts. Paleobiology, 25(1): 58-74. https://doi.org/10.1666/0094-8373(1999)025<0058:moic>2.3.co;2 doi: 10.1666/0094-8373(1999)025<0058:moic>2.3.co;2 |
| Du, Y. X., Bertinelli, A., Jin, X., et al., 2020. Integrated Conodont and Radiolarian Biostratigraphy of the Upper Norian in Baoshan Block, Southwestern China. Lethaia, 53(4): 1-13. https://doi.org/10.1111/let.12374 |
| Gabbott, S. E., Aldridge, R. J., Theron, J. N., 1995. A Giant Conodont with Preserved Muscle-Tissue from the Upper Ordovician of South-Africa. Nature, 374(6525): 800-803. https://doi.org/10.1038/374800a0 |
| Golding, M. L., 2018. Reconstruction of the Multielement Apparatus of Neogondolella ex gr. regalis Mosher, 1970 (Conodonta) from the Anisian (Middle Triassic) in British Columbia, Canada. Journal of Micropalaeontology, 37(1): 21-24. https://doi.org/10.5194/jm-37-21-2018 |
| Goudemand, N., Orchard, M. J., Urdy, S., et al., 2011. Synchrotron-Aided Reconstruction of the Conodont Feeding Apparatus and Implications for the Mouth of the First Vertebrates. PNAS, 108(21): 8720-8724. https://doi.org/10.1073/pnas.1101754108 |
| Goudemand, N., Orchard, M. J., Tafforeau, P., et al., 2012. Early Triassic Conodont Clusters from South China: Revision of the Architecture of the 15 Element Apparatuses of the Superfamily Gondolelloidea. Palaeontology, 55(5): 1021-1034. https://doi.org/10.1111/j.1475-4983.2012.01174.x |
| Huang, J. Y., Hu, S. X., Zhang, Q. Y., et al., 2019a. Gondolelloid Multielement Conodont Apparatus (Nicoraella) from the Middle Triassic of Yunnan Province, Southwestern China. Palaeogeography, Palaeoclimatology, Palaeoecology, 522: 98-110. https://doi.org/10.1016/j.palaeo.2018.07.015 |
| Huang, J. Y., Martínez-Pérez, C., Hu, S. X., et al., 2019b. Middle Triassic Conodont Apparatus Architecture Revealed by Synchrotron X-Ray Microtomography. Palaeoworld, 28(4): 429-440. https://doi.org/10.1016/j.palwor.2018.08.003 |
| Karádi, V., 2018. Middle Norian Conodonts from the Buda Hills, Hungary: an Exceptional Record from the Western Tethys. Journal of Iberian Geology, 44(1): 155-174. https://doi.org/10.1007/s41513-017-0009-3 |
| Karádi, V., Cau, A., Mazza, M., et al., 2020. The Last Phase of Conodont Evolution during the Late Triassic: Integrating Biostratigraphic and Phylogenetic Approaches. Palaeogeography, Palaeoclimatology, Palaeoecology, 549: 109144. https://doi.org/10.1016/j.palaeo.2019.03.045. |
| Koike, T., Yamakita, S., Kadota, N., 2004. A Natural Assemblage of Ellisonia sp. cf. E. triassica Müller (Vertebrata: Conodonta) from the Uppermost Permian in the Suzuka Mountains, Central Japan. Paleontological Research, 8(4): 241-253. https://doi.org/10.2517/prpsj.8.241 |
| Kolar-Jurkovšek, T., Dzicki, A. G. A., Jurkovšek, B., 2005. Conodonts and Foraminifera from the "Raibl Beds" (Carnian) of the Karavanke Mountains, Slovenia: Stratigraphical and Palaeobiological Implications. Geological Quarterly, 49(4): 429-438 http://www.researchgate.net/publication/286585232_Conodonts_and_foraminifera_from_the_Raibl_Beds_Carnian_of_the_Karavanke_Mountains_Slovenia_Stratigraphical_and_palaeobiological_implications |
| Kolar-Jurkovšek, T., Jurkovšek, B., 2010. New Paleontological Evidence of the Carnian Strata in the Mežica Area (Karavanke Mts, Slovenia): Conodont Data for the Carnian Pluvial Event. Palaeogeography, Palaeoclimatology, Palaeoecology, 290(1-4): 81-88. https://doi.org/10.1016/j.palaeo.2009.06.015 |
| Kolar-Jurkovšek, T., Martínez-Pérez, C., Jurkovšek, B., et al., 2018. New Clusters of Pseudofurnishius murcianus from the Middle Triassic of Slovenia (Dinarides). Bulletins of American Paleontology, 395/396: 149-163 http://www.researchgate.net/publication/328673314_New_clusters_of_Pseudofurnishius_murcianus_from_the_Middle_Triassic_of_Slovenia_Dinarides |
| Kozur, H., 1989. The Taxonomy of the Gondolellid Conodonts in the Permian and Triassic. Courier Forschungs Institut Senckenberg, 117: 409-469 http://www.researchgate.net/publication/284060687_The_taxonomy_of_the_gondolellid_conodont_in_the_Permian_and_Triassic |
| Kozur, H., 2003. Integrated Ammonoid, Conodont and Radiolarian Zonation of the Triassic and Some Remarks to Stage/Substage Subdivision and the Numeric Age of the Triassic Stages. Albertiana, 28: 57-74 http://www.researchgate.net/publication/286181376_Integrated_ammonoid_conodont_and_radiolarian_zonation_of_the_Triassic_and_some_remarks_to_StageSubstage_subdivision_and_the_numeric_age_of_the_Triassic_stages |
| Kozur, H., Mock, R., 1972. Neue Conodonten aus der Trias der Slowakei und ihre Straigraphische Bedeutung. Geologisch-Paläontologische Mitteilungen Innsbruck, 2: 1-20 http://www.researchgate.net/publication/285105149_Neue_Conodonten_aus_der_Trias_der_Slowakei_und_ihre_stratigraphische_Bedeutung |
| Kozur, H., Mock, R., 1991. New Middle Carnian and Rhaetian Conodonts from Hungary and the Alps. Stratigraphic Importance and Tectonic Implications for the Buda Mountains and Adjacent Areas. Jahrbuch der Geologischen Bundesanstalt, 134(2): 271-297 http://ci.nii.ac.jp/naid/10003918868 |
| Kozur, H., Mostler, H., 1970. Neue Conodonten aus der Trias. Ber. Nat-Med. Verein Innsbrunck, 58: 429-464 |
| Kozur, H., Mostler, H., 1972. Die Bedeutung der Conodonten für Stratigraphische und Paläogeographische Untersuchungen in der Trias. Mitt. Ges. Geol. Bergbaustud, 21: 777-311 http://www.researchgate.net/publication/285105326_Die_Bedeutung_der_Conodonten_fur_stratigraphische_und_palaogeographische_Untersuchungen_in_der_Trias |
| Lai, X. L., Mei, S. L., 2000. On Zonation and Evolution of Permian and Triassic Conodonts. Developments in Palaeontology and Stratigraphy, 18(C): 371-392. https://doi.org/10.1016/s0920-5446(00)80021-7 |
| Mastandrea, A., Ietto, F., Neri, C., et al., 1997. Conodont Biostratigraphy of Monte Cocuzzo Section (Upper Triassic, Catena Costiera-Calabria, Italy). Rivista Italiana di Paleontologia e Stratigrafia, 103(2): 173-182 http://www.researchgate.net/publication/287678683_Conodont_biostratigraphy_of_the_late_triassic_sequence_of_Monte_Cocuzzo_Catena_Costiera_Calabria_Italy |
| Mastandrea, A., Neri, C., Jetto, F., et al., 1999. Misikella ultima Kozur & Kock, 1991: First Evidence of Late Rhaetian Conodonts in Calabria (Southern Italy). Bollettino della Società Paleontologica Italiana, 37(2/3): 497-506 |
| Moix, P., Kozur, H. W., Stampfli, G. M., et al., 2007. New Paleontological, Biostratigraphical and Paleogeographic Results from the Triassic of the Mersin Mélange, SE Turkey. In: Lucas, S. G., Spielmann, J. A., eds., The Global Triassic. New Mexico Museum of Natural History and Science Bulletin, 41: 282-311 |
| Mosher, L. C., 1968. Triassic Conodonts from Western North America and Europe and Their Correlation. Journal of Paleontology, 42: 895-946 |
| Mosher, L. C., 1970. New Conodont Species as Triassic Guide Fossils. Journal of Paleontology, 44(4): 737-742 |
| Orchard, M. J., 1991. Upper Triassic Conodont Biochronology and New Index Species from the Canadian Cordillera. In: Orchard, M. J., McCracken, A. D., eds., Ordovician to Triassic Conodont Paleontology of the Canadian Cordillera. Geological Survey of Canada Bulletin, 417: 299-335 |
| Orchard, M. J., Rieber, H., 1999. Multielement Neogondolella (Conodonta, Upper Permian-Middle Triassic). Bollettino della Societa Paleontologica Italiana, 37(2/3): 475-488 http://ci.nii.ac.jp/naid/10020877422 |
| Orchard, M. J., 2005. Multielement Conodont Apparatuses of Triassic Gondolelloidea. Special Papers in Palaeontology, 73: 73-101 http://www.researchgate.net/publication/273694027_Multielement_Conodont_Apparatuses_of_Triassic_Gondolelloidea |
| Orchard, M. J., 2018. The Lower-Middle Norian (Upper Triassic) Boundary: New Conodont Taxa and a Refined Biozonation. Bulletins of American Paleontology, 395/396: 165-193 http://www.researchgate.net/publication/328673220_The_Lower-Middle_Norian_Upper_Triassic_boundary_New_conodont_taxa_and_a_refined_biozonation |
| Orchard, M. J., Zonneveld, J. P., Johns, M. J., et al., 2001. Fossil Succession and Sequence Stratigraphy of the Upper Triassic of Black Bear Ridge, Northeast British Columbia, a GSSP Prospect for the Carnian-Norian Boundary. Albertiana, 25: 10-22 http://www.researchgate.net/publication/263444703_Fossil_succession_and_sequence_stratigraphy_of_the_Upper_Triassic_of_Black_Bear_Ridge_northeast_British_Columbia_a_GSSP_prospect_for_the_Carnian-Norian_boundary |
| Purnell, M. A., 1993. Feeding Mechanisms in Conodonts and the Function of the Earliest Vertebrate Hard Tissues. Geology, 21(4): 375-377. https://doi.org/10.1130/0091-7613(1993)021<0375:fmicat>2.3.co;2 doi: 10.1130/0091-7613(1993)021<0375:fmicat>2.3.co;2 |
| Purnell, M. A., Aldridge, R. J., Donoghue, P. C., et al., 1995. Conodonts and the First Vertebrates. Endeavour, 19(1): 20-27. https://doi.org/10.1016/0160-9327(95)98890-r |
| Purnell, M. A., Donoghue, P. C. J., 1997. Architecture and Functional Morphology of the Skeletal Apparatus of Ozarkodinid Conodonts. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences, 352(1361): 1545-1564. https://doi.org/10.1098/rstb.1997.0141 |
| Purnell, M. A., Donoghue, P. C. J., 1998. Skeletal Architecture, Homologies and Taphonomy of Ozarkodinid Conodonts. Palaeontology, 41(1): 57-102 http://ci.nii.ac.jp/naid/10003918871 |
| Purnell, M. A., Donoghue, P. C. J., 2005. Between Death and Data: Biases in Interpretation of the Fossil Record of Conodonts. Special Papers in Palaeontology, 73: 7-25 http://www.researchgate.net/publication/228633068_Between_death_and_data_Biases_in_interpretation_of_the_fossil_record_of_conodonts |
| Purnell, M. A., Donoghue, P. C. J., Aldridge, R. J., 2000. Orientation and Anatomical Notation in Conodonts. Journal of Paleontology, 74(1): 113-122. https://doi.org/10.1666/0022-3360(2000)074<0113:oaanic>2.0.co;2 doi: 10.1666/0022-3360(2000)074<0113:oaanic>2.0.co;2 |
| Purnell, M. A., Zhang, M. H., Jiang, H. S., 2018. Reconstruction, Composition and Homology of Conodont Skeletons: A Response to Agematsu et al. Palaeontology, 61(5): 793-796. https://doi.org/10.1111/pala.12387 |
| Ramovš, A., 1977. Skelettapparat von Pseudofurnishius murcianus (Conodontophorida) im Mitteltrias Sloweniens (NW Jugoslawien). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 153: 361-399 http://www.researchgate.net/publication/285880207_Skelettapparat_von_Pseudofurnishius_murcianus_Conodontophorida_in_der_Mitteltrias_Sloweniens_NW_Jugoslawien |
| Ramovš, A., 1978. Mitteltriassische Conodonten-Clusters in Slowenien, NW Jugoslawien. Paläontologische Zeitschrift, 52(1/2): 129-137. https://doi.org/10.1007/bf03006734 |
| Rieber, H., 1980. Ein Conodonten-Cluster aus der Grenzbitumenzone (Mittlere Trias) des Monte San Giorgio (Kt. Tessin/Schweiz). Annalen des Naturhistorischen Museums Wien, 83: 265-274. https://www.jstor.org/stable/41768810 |
| Rigo, M., Mazza, M., Karádi, V., et al., 2018. New Upper Triassic Conodont Biozonation of the Tethyan Realm. In: Tanner, L. H., ed., The Late Triassic World: Earth in a Time of Transition. Springer, 189-235 |
| Roghi, G., Mietto, P., Dalla Vecchia, F. M., 1995. Contribution to the Conodont Biostratigraphy of the Dolomia di Forni (Upper Triassic, Carnia, NE Italy). Memorie di Scienze Geologische Universita di Padova, 47: 125-133 http://www.researchgate.net/publication/256103845_Contribution_to_the_Conodont_Biostratigraphy_of_the_Dolomia_di_Forni_Upper_Triassic_Carnia_NE_Italy |
| Ryley, C. C., Fåhraeus, L. E., 1994. Two New Genera Comperniodontella n. gen. and Galeodontella n. gen., and New Multielement Species of Chirodella HIRSCHMANN, 1959 and Cypridodella MOSHER, 1968 (Conodonta) from the Mamonia Complex. Neues Jahrbuch fur Geologie und Palaontologie-Abhandlungen, 193(1): 21-54 |
| Sun, Z. Y., Hao, W. C., Sun, Y. L., et al., 2009. The Conodont Genus Nicoraella and a New Species from the Anisian of Guizhou, South China. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 252(2): 227-235. https://doi.org/10.1127/0077-7749/2009/0252-0227 |
| Sun, Z. Y., Liu, S., Ji, C., et al., 2020a. Synchrotron-Aided Reconstruction of the Prioniodinin Multielement Conodont Apparatus (Hadrodontina) from the Lower Triassic of China. Palaeogeography, Palaeoclimatology, Palaeoecology, 560: 109913. https://doi.org/10.1016/j.palaeo.2020.109913 |
| Sun, Z. Y., Liu, S., Ji, C., et al., 2020b. Gondolelloid Multielement Conodont Apparatus (Scythogondolella) from the Lower Triassic of Jiangsu, East China, Revealed by High-Resolution X-Ray Microtomography. Palaeoworld. https://doi.org/10.1016/j.palwor.2020.06.001 |
| Takahashi, S., Yamakita, S., Suzuki, N., 2019. Natural Assemblages of the Conodont Clarkina in Lowermost Triassic Deep-Sea Black Claystone from Northeastern Japan, with Probable Soft-Tissue Impressions. Palaeogeography, Palaeoclimatology, Palaeoecology, 524: 212-229. https://doi.org/10.1016/j.palaeo.2019.03.034 |
| Tolmacheva, T. Y., Purnell, M. A., 2002. Apparatus Composition, Growth, and Survivorship of the Lower Ordovician Conodont Paracordylodus gracilis Lindström, 1955. Palaeontology, 45(2): 209-228. https://doi.org/10.1111/1475-4983.00234 |
| Wang, X. D., Du, Y. X., Shi, Z. Q., et al., 2019. Characteristics of Conodonts and Sedimentary Environment of Upper Norian Nanshuba and Dashuitang Formations in Baoshan, Yunnan Province. Acta Palaeontologica Sinica, 58(1): 79-91 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTotal-GSWX201901006.htm |
| Wang, Z. H., Dong, Z. Z., 1985. Discovery of Conodont Epigondolella Fauna from Late Triassic in Baoshan Area, Western Yunnan. Acta Micropalaeontologica Sinica, 2: 125-132 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-WSGT198502002.htm |
| Yuan, J. L., Jiang, H. S., Wang, D. C., 2015. LST: A New Inorganic Heavy Liquid Used in Conodont Separation. Geological Science and Technology Information, 34(5): 225-230 (in Chinese with English Abstract) http://www.en.cnki.com.cn/Article_en/CJFDTotal-DZKQ201505035.htm |
| Zhang, M. H., Jiang, H. S., Purnell, M. A., et al., 2017. Testing Hypotheses of Element Loss and Instability in the Apparatus Composition of Complex Conodonts: Articulated Skeletons of Hindeodus. Palaeontology, 60(4): 595-608. https://doi.org/10.1111/pala.12305 |
| Zhang, Y. Z., Zhang, D. H., Liu, S. R., et al., 1996. Lithostratigraphy of Yunnan Province. China University of Geosciences Press, Wuhan. 366 (in Chinese) |
| Zhao, Y. J., Zheng, C., Huang, L., et al., 2012. Catastrophic Event Characteristics of Stratigraphic Slope Slumping Accumulation: A Case Study of the Upper Triassic Nanshuba Formation in Baoshan Area, Western Yunnan Province. Geological Bulletin of China, 31(5): 745-757 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-ZQYD201205012.htm |

Figures(7) / Tables(2)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: