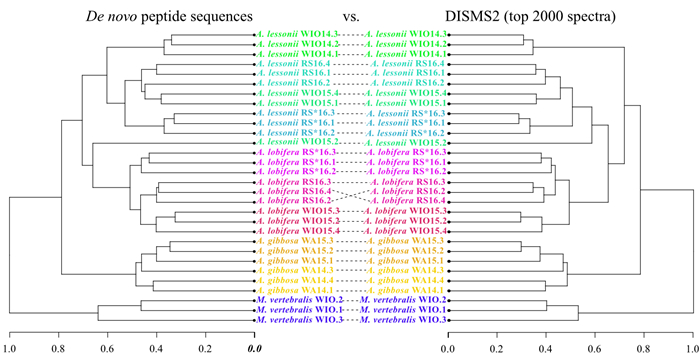
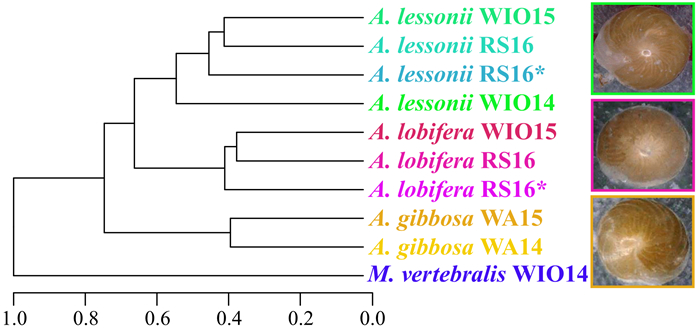
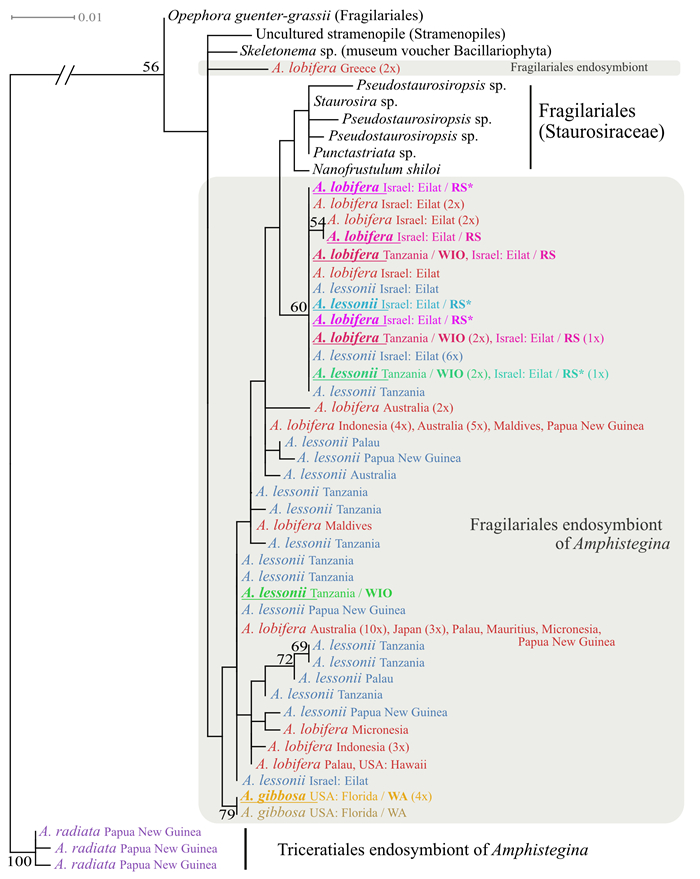
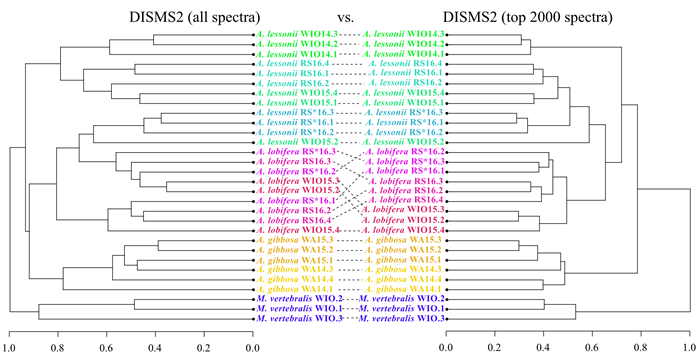
| Citation: | Marleen Stuhr, Bernhard Blank-Landeshammer, Achim Meyer, Vera Baumeister, Jörg Rahnenführer, Albert Sickmann, Hildegard Westphal. Proteome-Based Clustering Approaches Reveal Phylogenetic Insights into Amphistegina. Journal of Earth Science, 2022, 33(6): 1469-1479. doi: 10.1007/s12583-022-1609-1

|
Foraminifera are highly diverse and have a long evolutionary history. As key bioindicators, their phylogenetic schemes are of great importance for paleogeographic applications, but may be hard to recognize correctly. The phylogenetic relationships within the prominent genus

| Aebersold, R., Mann, M., 2003. Mass Spectrometry-Based Proteomics. Nature, 422(6928): 198–207. https://doi.org/10.1038/nature01511 |
| Alves, B. M., Júnior, M. N., 2020. Foraminiferal Assemblage Structure from Brazilian Tropical Urbanized Beaches (~7°S). Anais da Academia Brasileira de Ciências, 92(2): 1–15. https://doi.org/10.1590/0001-3765202020190486 |
| Baer, B., Millar, A. H., 2016. Proteomics in Evolutionary Ecology. Journal of Proteomics, 135: 4–11. https://doi.org/10.1016/j.jprot.2015.09.031 |
| Barnes, K. H., 2016. Diversity and Distribution of Diatom Endosymbionts in Amphistegina spp. (Foraminifera) Based on Molecular and Morphological Techniques: [Dissertation]. University of South Florida, St. Petersburg, Florida. 160 |
| Bentov, S., Brownlee, C., Erez, J., 2009. The Role of Seawater Endocytosis in the Biomineralization Process in Calcareous Foraminifera. Proceedings of the National Academy of Sciences of the United States of America, 106(51): 21500–21504. https://doi.org/10.1073/pnas. 0906636106 doi: 10.1073/pnas.0906636106 |
| Bhatt, K. A., Trivedi, M. H., 2018. Molecular Studies on Foraminifers: Past, Present, and Future. Journal of Foraminiferal Research, 48(3): 193–209. https://doi.org/10.2113/gsjfr.48.3.193 |
| Blank-Landeshammer, B., Kollipara, L., Biß, K., et al., 2017. Combining de Novo Peptide Sequencing Algorithms, a Synergistic Approach to Boost both Identifications and Confidence in Bottom-up Proteomics. Journal of Proteome Research, 16(9): 3209–3218. https://doi.org/10.1021/acs.jproteome.7b00198 |
| Boudagher-Fadel, M. K., 2018. Evolution and Geological Significance of Larger Benthic Foraminifera. Second Edition. UCL Press, London. 693. https://doi.org/10.2307/j.ctvqhsq3 |
| Carnahan, E. A., Hoare, A. M., Hallock, P., et al., 2009. Foraminiferal Assemblages in Biscayne Bay, Florida, USA: Responses to Urban and Agricultural Influence in a Subtropical Estuary. Marine Pollution Bulletin, 59(8–12): 221–233. https://doi.org/10.1016/j.marpolbul.2009.08.008 |
| Chambers, M. C., Maclean, B., Burke, R., et al., 2012. A Cross-Platform Toolkit for Mass Spectrometry and Proteomics. Nature Biotechnology, 30(10): 918–920. https://doi.org/10.1038/nbt.2377 |
| d'Orbigny, A. D., 1826. Tableau Méthodique de la Classe des Céphalopodes. Annales des Sciences Naturelles, 7: 96–314 |
| Debenay, J. -P., 2013. A Guide to 1 000 Foraminifera from Southwestern Pacific, New Caledonia. Muséum National d'Histoire Naturelle, Paris. 384 |
| Dice, L., 1945. Measures of the Amount of Ecologic Association between Species. Ecology, 26(3): 297–302. https://doi.org/10.2307/1932409 |
| Doo, S. S., Hamylton, S., Byrne, M., 2012. Reef-Scale Assessment of Intertidal Large Benthic Foraminifera Populations on one Tree Island, Great Barrier Reef and Their Future Carbonate Production Potential in a Warming Ocean. Zoological Studies, 51(8): 1298–1307. https://doi.org/10.1007/s00435-012-0164-1 |
| Ehrenberg, C. G., 1838. Über die Bildung der Kreidefelsen und des Kreidemergels Durch Unsichtbare Organismen. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin, Berlin. 59–147 |
| Emrich, K., Martinez-Colon, M., Alegria, H., 2017. Is Untreated Sewage Impacting Coral Reefs of Caye Caulker, Belize? The Journal of Foraminiferal Research, 47(1): 20–33. https://doi.org/10.2113/gsjfr.47.1.20 |
| Flakowski, J., Bolivar, I., Fahrni, J., et al., 2005. Actin Phylogeny of Foraminifera. Journal of Foraminiferal Research, 35(2): 93–102. https://doi.org/10.2113/35.2.93 |
| Förderer, M., Langer, M. R., 2019. Exceptionally Species-Rich Assemblages of Modern Larger Benthic Foraminifera from Nearshore Reefs in Northern Palawan (Philippines). Revue de Micropaléontologie, 65: 100387. https://doi.org/10.1016/j.revmic.2019.100387 |
| Förderer, M., Rödder, D., Langer, M. R., 2018. Patterns of Species Richness and the Center of Diversity in Modern Indo-Pacific Larger Foraminifera. Scientific Reports, 8(1): 8189. https://doi.org/10.1038/s41598-018-26598-9 |
| Gibb, S., 2015. readMzXmlData: Reads Mass Spectrometry Data in mzXML Format. [2021-11-10]. https://github.com/sgibb/readMzXmlData/ |
| Glas, M. S., Fabricius, K. E., de Beer, D., et al., 2012. The O2, pH and Ca2+ Microenvironment of Benthic Foraminifera in a High CO2 World. PLoS One, 7(11): e50010. https://doi.org/10.1371/journal.pone.0050010 |
| Glez-Peña, D., Gómez-Blanco, D., Reboiro-Jato, M., et al., 2010. ALTER: Program-Oriented Conversion of DNA and Protein Alignments. Nucleic Acids Research, 38: W14–W18. https://doi.org/10.1093/nar/gkq321 |
| Green, M. R., Sambrook, J., 2012. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y. 2028 |
| Guastella, R., Marchini, A., Caruso, A., et al., 2019. "Hidden Invaders" Conquer the Sicily Channel and Knock on the Door of the Western Mediterranean Sea. Estuarine, Coastal and Shelf Science, 225: 106234. https://doi.org/10.1016/j.ecss.2019.05.016 |
| Hallock, P., 1985. Why are Larger Foraminifera Large? Paleobiology, 11(2): 195–208. https://doi.org/10.1017/s0094837300011507 |
| Hallock, P., 1999. Symbiont-Bearing Foraminifera. In: Sen Gupta, B. K., ed., Modern Foraminifera. Kluwer Academic Publishers. 123–368 |
| Hallock, P., Forward, L. B., Hansen, H. J., 1986. Influence of Environment on the Test Shape of Amphistegina. The Journal of Foraminiferal Research, 16(3): 224–231. https://doi.org/10.2113/gsjfr.16.3.224 |
| Hallock, P., Glenn, E. C., 1986. Larger Foraminifera: A Tool for Paleoenvironmental Analysis of Cenozoic Carbonate Depositional Facies. PALAIOS, 1(1): 55–64. https://doi.org/10.2307/3514459 |
| Hallock, P., Hansen, J., 1979. Depth Adaptation in Amphistegina: Change in Lamellar Thickness. Bulletin of the Geological Society of Denmark, 27: 99–104. https://doi.org/10.37570/bgsd-1978-27-10 |
| Hallock, P., Lidz, B. H., Cockey-Burkhard, E. M., et al., 2003. Foraminifera as Bioindicators in Coral Reef Assessment and Monitoring: The Foram Index. Environmental Monitoring and Assessment, 81(1/2/3): 221–238. https://doi.org/10.1023/a:1021337310386 |
| Hallock, P., Talge, H. K., Smith, K., et al., 1993. Bleaching in a Reef-Dwelling Foraminifer, Amphistengina Gibbosa. Proceedings of the 7th International Coral Reef Symposium, 1: 44–49 |
| Hansen, H. J., Buchard, B., 1977. Depth Distribution of Amphistegina in the Gulf of Elat, Israel. Utrecht Micropaleontological Bulletins, 1977: 205–224 |
| Haynes, J. R., 1992. Supposed Pronounced Ecophenotypy in Foraminifera. Journal of Micropalaeontology, 11(1): 59–63. https://doi.org/10.1144/jm.11.1.59 |
| Heck, M., Neely, B. A., 2020. Proteomics in Non-Model Organisms: A New Analytical Frontier. Journal of Proteome Research, 19(9): 3595–3606. https://doi.org/10.1021/acs.jproteome.0c00448 |
| Hohenegger, J., 2014. Species as the Basic Units in Evolution and Biodiversity: Recognition of Species in the Recent and Geological Past as Exemplified by Larger Foraminifera. Gondwana Research, 25(2): 707–728. https://doi.org/10.1016/j.gr.2013.09.009 |
| Hohenegger, J., 2004. Depth Coenoclines and Environmental Considerations of Western Pacific Larger Foraminifera. The Journal of Foraminiferal Research, 34(1): 9–33. https://doi.org/10.2113/0340009 |
| Hottinger, L., Halicz, E., Reiss, Z., 1993. Recent Foraminiferida from the Gulf of Aqaba, Red Sea. Slovenian Academy of Sciences and Arts and Swiss Academy of Natural Sciences, Ljubljana. 181 |
| Kozlov, A. M., Darriba, D., Flouri, T., et al., 2019. RAxML-NG: A Fast, Scalable and User-Friendly Tool for Maximum Likelihood Phylogenetic Inference. Bioinformatics, 35(21): 4453–4455. https://doi.org/10.1093/bioinformatics/btz305 |
| Kumar, S., Stecher, G., Li, M., et al., 2018. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Molecular Biology and Evolution, 35(6): 1547–1549. https://doi.org/10.1093/molbev/msy096 |
| Langer, M., 2008. Foraminifera from the Mediterranean and the Red Sea. In: Aqaba-Eilat, the Improbable Gulf: Environment, Biodiversity and Preservation. Magnes Press, Jerusalem. 397–415 |
| Langer, M. R., Hottinger, L., 2000. Biogeography of Selected "Larger" Foraminifera. Micropaleontology, 46: 105–126 |
| Langer, M. R., Silk, M. T., Lipps, J. H., 1997. Global Ocean Carbonate and Carbon Dioxide Production: The Role of Reef Foraminifera. The Journal of Foraminiferal Research, 27(4): 271–277. https://doi.org/10.2113/gsjfr.27.4.271 |
| Langer, M. R., Weinmann, A. E., Lötters, S., et al., 2013. Climate-Driven Range Extension of Amphistegina (Protista, Foraminiferida): Models of Current and Predicted Future Ranges. PLoS One, 8(2): e54443. https://doi.org/10.1371/journal.pone.0054443 |
| Larsen, A. R., 1978. Phylogenetic and Paleobiogeographical Trends in the Foraminiferal genus Amphistegina. Revista Española de Micropaleontología, X: 217–243 |
| Lee, J., 2006. Algal Symbiosis in Larger Foraminifera. Symbiosis, 42(2): 63–75 |
| Leutenegger, S., 1977. Ultrastructure de Foraminifères Perforés et Imperforés Ainsi Que Leurs Symbiotes. Cahiers de Micropaléontologie, 3: 5–7 |
| Loeblich, A. R. Jr., Tappan, H., 1988. Foraminiferal Genera and Their Classification, Vol. 1. Van Nostrand Reinhold. 2115 |
| Loeblich, A. R. Jr., Tappan, H., 1994. Foraminifera of the Sahul Shelf and Timor Sea. Cushman Foundation Special Publications No. 31. 666 |
| Loeblich, A. R. Jr., Tappan, H., Beckmann, J. P., et al., 1957. Studies in Foraminifera. United States National Museum Bulletin, 215: 412 |
| MacCoss, M. J., 2005. Computational Analysis of Shotgun Proteomics Data. Current Opinion in Chemical Biology, 9(1): 88–94. https://doi.org/10.1016/j.cbpa.2004.12.010 |
| Macher, J. N., Prazeres, M., Taudien, S., et al., 2021. Integrating Morphology and Metagenomics to Understand Taxonomic Variability of Amphisorus (Foraminifera, Miliolida) from Western Australia and Indonesia. PLoS One, 16(1): e0244616. https://doi.org/10.1371/journal.pone.0244616 |
| Morard, R., Füllberg, A., Brummer, G. J. A., et al., 2019. Genetic and Morphological Divergence in the Warm-Water Planktonic Foraminifera Genus Globigerinoides. PLoS One, 14(12): e0225246. https://doi.org/10.1371/journal.pone.0225246 |
| Mouanga, G. H., Langer, M. R., 2014. At the Front of Expanding Ranges: Shifting Community Structures at Amphisteginid Species Range Margins in the Mediterranean Sea. Neues Jahrbuch Für Geologie Und Paläontologie-Abhandlungen, 271(2): 141–150. https://doi.org/10.1127/0077-7749/2014/0381 |
| Murray, J., 2006. Ecology and Applications of Benthic Foraminifera. Palaeogeography, Palaeoclimatology, Palaeoecology, 95: 1–426. https://doi.org/10.1017/cbo9780511535529 |
| Narayan, G. R., Reymond, C. E., Stuhr, M., et al., 2022. Response of Large Benthic Foraminifera to Climate and Local Changes: Implications for Future Carbonate Production. Sedimentology, 69(1): 121–161. https://doi.org/10.1111/sed.12858 |
| Narayan, Y. R., Lybolt, M., Zhao, J. X., et al., 2015. Holocene Benthic Foraminiferal Assemblages Indicate Long-Term Marginality of Reef Habitats from Moreton Bay, Australia. Palaeogeography, Palaeoclimatology, Palaeoecology, 420: 49–64. https://doi.org/10.1016/j.palaeo.2014.12.010 |
| Palmblad, M., Deelder, A. M., 2012. Molecular Phylogenetics by Direct Comparison of Tandem Mass Spectra. Rapid Communications in Mass Spectrometry, 26(7): 728–732. https://doi.org/10.1002/rcm.6162 |
| Paradis, E., Claude, J., Strimmer, K., 2004. APE: Analyses of Phylogenetics and Evolution in R Language. Bioinformatics, 20(2): 289–290. https://doi.org/10.1093/bioinformatics/btg412 |
| Pawlowski, J., Holzmann, M., 2008. Diversity and Geographic Distribution of Benthic Foraminifera: A Molecular Perspective. Biodiversity and Conservation, 17(2): 317–328. https://doi.org/10.1007/s10531-007-9253-8 |
| Pawlowski, J., Holzmann, M., 2002. Molecular Phylogeny of Foraminifera a Review. European Journal of Protistology, 38(1): 1–10. https://doi.org/10.1078/0932-4739-00857 |
| Pawlowski, J., Holzmann, M., Tyszka, J., 2013. New Supraordinal Classification of Foraminifera: Molecules Meet Morphology. Marine Micropaleontology, 100: 1–10. https://doi.org/10.1016/j.marmicro.2013. 04.002 doi: 10.1016/j.marmicro.2013.04.002 |
| Pawlowski, J., Lecroq, B., 2010. Short rDNA Barcodes for Species Identification in Foraminifera. Journal of Eukaryotic Microbiology, 57(2): 197–205. https://doi.org/10.1111/j.1550-7408.2009.00468.x |
| Pawlowski, J., Lejzerowicz, F., Esling, P., 2014. Next-Generation Environmental Diversity Surveys of Foraminifera: Preparing the Future. The Biological Bulletin, 227(2): 93–106. https://doi.org/10.1086/bblv227n2p93 |
| Pisapia, C., Kateb, A. E., Hallock, P., et al., 2017. Assessing Coral Reef Health in the North Ari Atoll (Maldives) Using the FoRAM Index. Marine Micropaleontology, 133: 50–57. https://doi.org/10.1016/j.marmicro.2017.06.001 |
| Pochon, X., Montoya-Burgos, J. I., Stadelmann, B., et al., 2006. Molecular Phylogeny, Evolutionary Rates, and Divergence Timing of the Symbiotic Dinoflagellate Genus Symbiodinium. Molecular Phylogenetics and Evolution, 38(1): 20–30. https://doi.org/10.1016/j.ympev.2005.04.028 |
| Pomar, L., Hallock, P., 2008. Carbonate Factories: A Conundrum in Sedimentary Geology. Earth-Science Reviews, 87(3/4): 134–169. https://doi.org/10.1016/j.earscirev.2007.12.002 |
| Prazeres, M., Martínez-Colón, M., Hallock, P., 2020a. Foraminifera as Bioindicators of Water Quality: The FoRAM Index Revisited. Environmental Pollution, 257: 113612. https://doi.org/10.1016/j.envpol.2019.113612 |
| Prazeres, M., Morard, R., Roberts, T. E., et al., 2020b. High Dispersal Capacity and Biogeographic Breaks Shape the Genetic Diversity of a Globally Distributed Reef-Dwelling Calcifier. Ecology and Evolution, 10(12): 5976–5989. https://doi.org/10.1002/ece3.6335 |
| Prazeres, M., Renema, W., 2019. Evolutionary Significance of the Microbial Assemblages of Large Benthic Foraminifera. Biological Reviews, 94(3): 828–848. https://doi.org/10.1111/brv.12482 |
| Prazeres, M., Roberts, T. E., Ramadhani, S. F., et al., 2021. Diversity and Flexibility of Algal Symbiont Community in Globally Distributed Larger Benthic Foraminifera of the Genus Amphistegina. BMC Microbiology, 21(1): 243. https://doi.org/10.1186/s12866-021-02299-8 |
| Prazeres, M., Martins, S. E., Bianchini, A., 2011. Biomarkers Response to Zinc Exposure in the Symbiont-Bearing Foraminifer Amphistegina Lessonii (Amphisteginidae, Foraminifera). Journal of Experimental Marine Biology and Ecology, 407(1): 116–121. https://doi.org/10.1016/j.jembe.2011.07.015 |
| Prazeres, M., Martins, S. E., Bianchini, A., 2012. Assessment of Water Quality in Coastal Waters of Fernando de Noronha, Brazil: Biomarker Analyses in Amphistegina Lessonii. The Journal of Foraminiferal Research, 42(1): 56–65. https://doi.org/10.2113/gsjfr.42.1.56 |
| Prazeres, M., Uthicke, S., Pandolfi, J. M., 2015. Ocean Acidification Induces Biochemical and Morphological Changes in the Calcification Process of Large Benthic Foraminifera. Proceedings Biological Sciences, 282: 8–11. https://doi.org/10.1098/rspb.2014.2782 |
| Ramadhani, S. F., Roberts, T. E., Stuhr, M., et al., 2022. Patterns of Host-Symbiont Genetic Diversity and Specificity in the Reef-Dwelling Large Benthic Foraminifera Amphistegina lessonii. Coral Reefs, Under Review |
| Renema, W., 2003. Larger Foraminifera on Reefs around Bali (Indonesia). Zool Verh Leiden, 345: 337–366 |
| Renema, W., 2018. Terrestrial Influence as a Key Driver of Spatial Variability in Large Benthic Foraminiferal Assemblage Composition in the Central Indo-Pacific. Earth-Science Reviews, 177: 514–544. https://doi.org/10.1016/j.earscirev.2017.12.013 |
| Renema, W., 2019. Large Benthic Foraminifera in Low-Light Environments. Mesophotic Coral Ecosystems, 553–562. https://doi.org/10.1007/978-3-319-92735-0_31 |
| Revell, L. J., 2012. Phytools: An R Package for Phylogenetic Comparative Biology (and other Things). Methods in Ecology and Evolution, 3(2): 217–223. https://doi.org/10.1111/j.2041-210X.2011.00169.x |
| Rieder, V., Blank-Landeshammer, B., Stuhr, M., et al., 2017a. DISMS2: A Flexible Algorithm for Direct Proteome-Wide Distance Calculation of LC-MS/MS Runs. BMC Bioinformatics, 18(1): 148. https://doi.org/10.1186/s12859-017-1514-2 |
| Rieder, V., Schork, K. U., Kerschke, L., et al., 2017b. Comparison and Evaluation of Clustering Algorithms for Tandem Mass Spectra. Journal of Proteome Research, 16(11): 4035–4044. https://doi.org/10.1021/acs.jproteome.7b00427 |
| Ross, B. J., Hallock, P., 2014. Chemical Toxicity on Coral Reefs: Bioassay Protocols Utilizing Benthic Foraminifers. Journal of Experimental Marine Biology and Ecology, 457: 226–235. https://doi.org/10.1016/j.jembe.2014.04.020 |
| Ross, B. J., Hallock, P., 2018. Challenges in Using Cell Tracker Green on Foraminifers that Host Algal Endosymbionts. PeerJ, 6: e5304. https://doi.org/10.7717/peerj.5304 |
| Schmidt, C., Heinz, P., Kucera, M., et al., 2011. Temperature-Induced Stress Leads to Bleaching in Larger Benthic Foraminifera Hosting Endosymbiotic Diatoms. Limnology and Oceanography, 56(5): 1587–1602. https://doi.org/10.4319/lo.2011.56.5.1587 |
| Schmidt, C., Morard, R., Almogi-Labin, A., et al., 2015. Recent Invasion of the Symbiont-Bearing Foraminifera Pararotalia into the Eastern Mediterranean Facilitated by the Ongoing Warming Trend. PLoS One, 10(8): e0132917. https://doi.org/10.1371/journal.pone.0132917 |
| Schmidt, C., Morard, R., Prazeres, M., et al., 2016. Retention of High Thermal Tolerance in the Invasive Foraminifera Amphistegina Lobifera from the Eastern Mediterranean and the Gulf of Aqaba. Marine Biology, 163(11): 1–13. https://doi.org/10.1007/s00227-016-2998-4 |
| Sørensen, T., 1948. Method of Establishing Groups of Equal Amplitude in Plant Sociology Based on Similarity of Species Content and Its Application to Analyses of the Vegetation on Danish Commons. Kongelige Danske Videnskabernes Selskab, Biologiske Skrifter, 5: 1–34 |
| Stainbank, S., Spezzaferri, S., Beccari, V., et al., 2020. Photic Stress on Coral Reefs in the Maldives: The Amphistegina Bleaching Index. Ecological Indicators, 113: 106257. https://doi.org/10.1016/j.ecolind.2020.106257 |
| Stuhr, M., Blank-Landeshammer, B., Reymond, C. E., et al., 2018a. Disentangling Thermal Stress Responses in a Reef-Calcifier and Its Photosymbionts by Shotgun Proteomics. Scientific Reports, 8: 3524. https://doi.org/10.1038/s41598-018-21875-z |
| Stuhr, M., Meyer, A., Reymond, C. E., et al., 2018b. Variable Thermal Stress Tolerance of the Reef-Associated Symbiont-Bearing Foraminifera Amphistegina Linked to Differences in Symbiont Type. Coral Reefs, 37(3): 811–824. https://doi.org/10.1007/s00338-018-1707-9 |
| Stuhr, M., Cameron, L. P., Blank-Landeshammer, B., et al., 2021. Divergent Proteomic Responses Offer Insights into Resistant Physiological Responses of a Reef-Foraminifera to Climate Change Scenarios. Oceans, 2(2): 281–314. https://doi.org/10.3390/oceans2020017 |
| Stuhr, M., Reymond, C. E., Rieder, V., et al., 2017. Reef Calcifiers are Adapted to Episodic Heat Stress but Vulnerable to Sustained Warming. PLoS One, 12(7): e0179753.https://doi.org/10.1371/journal.pone. 0179753 doi: 10.1371/journal.pone.0179753 |
| Stulpinaite, R., Hyams-Kaphzan, O., Langer, M. R., 2020. Alien and Cryptogenic Foraminifera in the Mediterranean Sea: A Revision of Taxa as Part of the EU 2020 Marine Strategy Framework Directive. Mediterranean Marine Science, 21: 719–758. https://doi.org/10.12681/mms.24673 |
| Talge, H. K., Hallock, P., 1995. Cytological Examination of Symbiont Loss in a Benthic Foraminifera, Amphistegina Gibbosa. Marine Micropaleontology, 26(1/2/3/4): 107–113. https://doi.org/10.1016/0377-8398(95)00015-1 |
| ter Kuile, B., Erez, J., 1984. In situ Growth Rate Experiments on the Symbiont-Bearing Foraminifera Amphistegina Lobifera and Amphisorus Hemprichii. The Journal of Foraminiferal Research, 14(4): 262–276. https://doi.org/10.2113/gsjfr.14.4.262 |
| ter Kuile, B., Erez, J., Padan, E., 1989. Mechanisms for the Uptake of Inorganic Carbon by Two Species of Symbiont-Bearing Foraminifera. Marine Biology, 103(2): 241–251. https://doi.org/10.1007/bf00543354 |
| Titelboim, D., Sadekov, A., Blumenfeld, M., et al., 2021. Monitoring of Heavy Metals in Seawater Using Single Chamber Foraminiferal Sclerochronology. Ecological Indicators, 120: 106931. https://doi.org/10.1016/j.ecolind.2020.106931 |
| Todd, R., 1976. Some Observations of Amphistegina (Foraminifera). Progress in Micropaleontology, Micropaleontology Press, New York. 382–394 |
| Triantaphyllou, M. V., Koukousioura, O., Dimiza, M. D., 2009. The Presence of the Indo-Pacific Symbiont-Bearing Foraminifer Amphistegina Lobifera in Greek Coastal Ecosystems (Aegean Sea, Eastern Mediterranean). Mediterranean Marine Science, 10(2): 73–86. https://doi.org/10.12681/mms.111 |
| Weber, A. A. T., Pawlowski, J., 2014. Wide Occurrence of SSU rDNA Intragenomic Polymorphism in Foraminifera and Its Implications for Molecular Species Identification. Protist, 165(5): 645–661. https://doi.org/10.1016/j.protis.2014.07.006 |
| Weinmann, A. E., Rödder, D., Lötters, S., et al., 2013. Heading for New Shores: Projecting Marine Distribution Ranges of Selected Larger Foraminifera. PLoS One, 8(4): e62182. https://doi.org/10.1371/journal.pone.0062182 |
| Zmiri, A., Kahan, D., Hochstein, S., et al., 1974. Phototaxis and Thermotaxis in some Species of Amphistegina (Foraminifera). The Journal of Protozoology, 21(1): 133–138. https://doi.org/10.1111/j.1550-7408.1974.tb03626.x |
Figures(4) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: