| Citation: | Peter Lunt, Xiwu Luan. SE Asian Cenozoic Larger Foraminifera: Taxonomic Questions, Apparent Radiation and Abrupt Extinctions. Journal of Earth Science, 2022, 33(6): 1378-1399. doi: 10.1007/s12583-022-1614-4

|
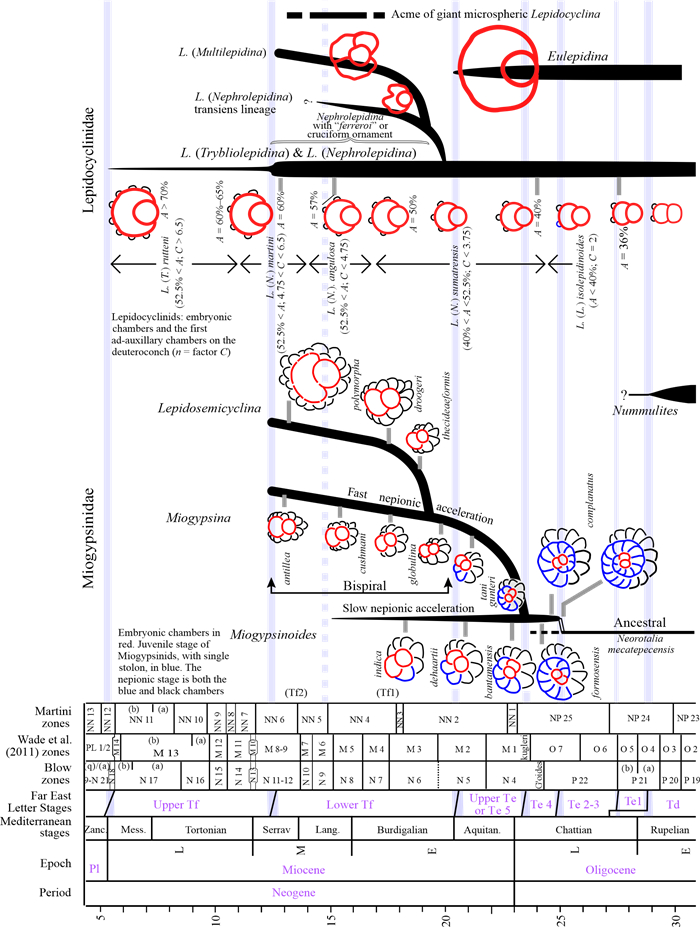
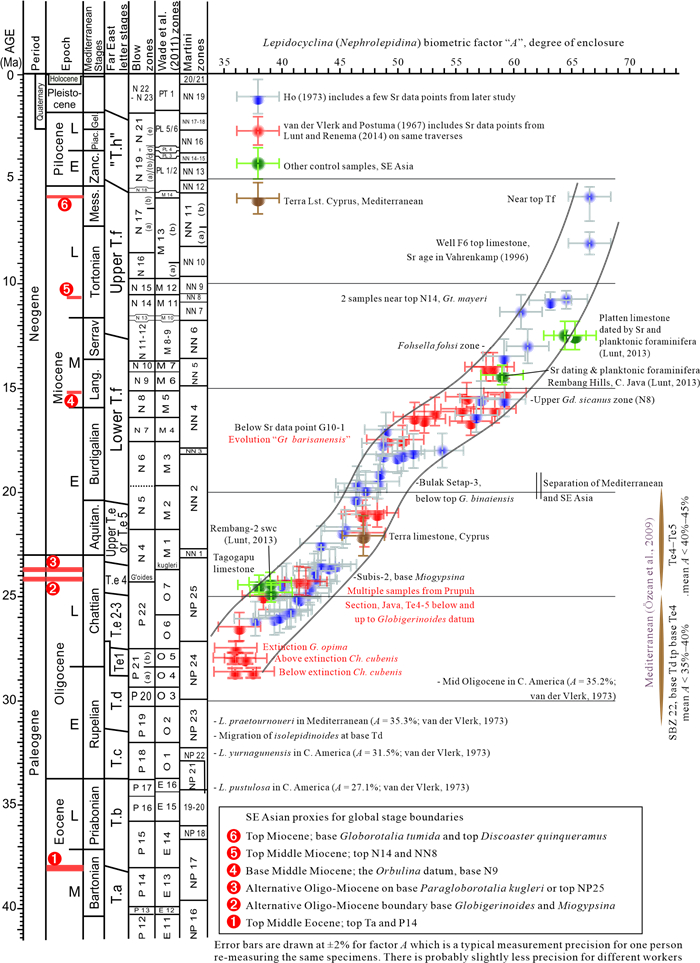
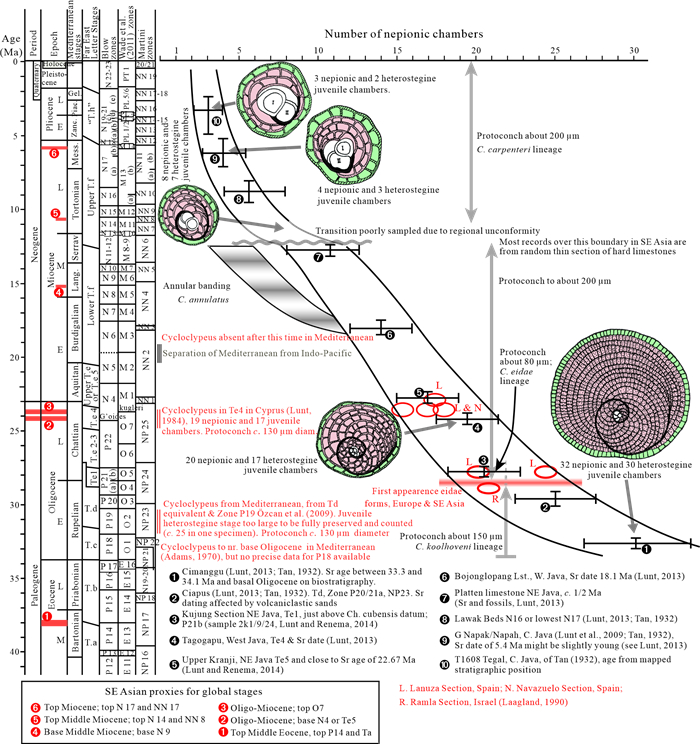
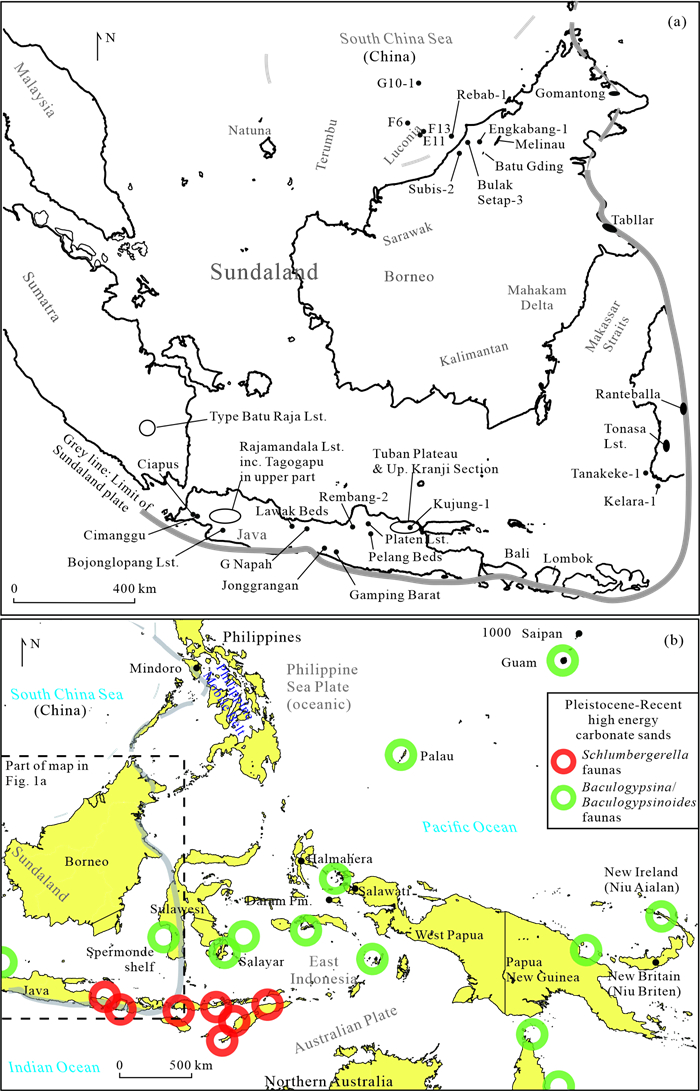
The Cenozoic larger foraminifera are important for biostratigraphy in SE Asia. This review examines the taxonomic inconsistencies of this group and especially the confusion over concepts of evolution, migration, radiation and extinction. It is proposed that from the Mediterranean to Indo-Pacific, Latest Eocene through Miocene, larger foraminifera populations were more homogenous than previously believed. Lineages show a slow morphological radiation interrupted by several extinctions. This pattern is superimposed on a long-term decline in larger foraminiferal abundance. The dating of the major biostratigraphic events are qualified. The main lineages of larger foraminifera can be classified by their evolutionary style. The classically "large" genera have highly peramorphic trends achieved through strong orthoselection, and these lineages were the most severely hit by a series of Mid-Eocene to Mid-Miocene extinction events. Other carbonate facies taxa that are traditionally grouped with the larger foraminifera are characterised by weak paramorphism, and these were much less affected by the extinction events. Some of these weakly peramorphic forms underwent Latest Miocene to recent radiation to and locally become rock-forming organisms. The three major and one minor larger foraminiferal extinction events since the Mid Eocene coincide with abrupt tectonic events across SE Asia. However, there are probably multiple causes for these extinctions, including global climatic changes.

| Abdulsamad, E. O., Barbieri, R., 1999. Foraminiferal Distribution and Palaeoecological Interpretation of the Eocene–Miocene Carbonates at al Jabal al Akhdar (Northeast Libya). Journal of Micropalaeontology, 18(1): 45–65. https://doi.org/10.1144/jm.18.1.45 |
| Abdulsamad, E. O., Bu-Argoub, F. M., Tmalla, A. F. A., 2009. A Stratigraphic Review of the Eocene to Miocene Rock Units in the al Jabal al Akhdar, NE Libya. Marine and Petroleum Geology, 26(7): 1228–1239. https://doi.org/10.1016/j.marpetgeo.2008.06.003 |
| Adams, C. G., 1965. The Foraminifera and Stratigraphy of the Melinau Limestone, Sarawak, and Its Importance in Tertiary Correlation. Quarterly Journal of the Geological Society, 121(1/2/3/4): 283–338. https://doi.org/10.1144/gsjgs.121.1.0283 |
| Adams, C. G., 1970. A Reconsideration of the East Indian Letter Classification of the Tertiary. Bulletins of the British Museum (Natural History), 19(3): 1–137 |
| Adams, C. G., 1983. Speciation, Phylogenesis, Tectonism, Climate and Eustasy: Factors in the Evolution of Cenozoic Larger Foraminifera. In: Sims, R. W., Price, J. H., Whalley, P. E. S., eds., Evolution, Time and Space: The Emergence of the Biosphere, Academic Press, London, New York. 255–289 |
| Adams, C. G., 1984. Neogene Larger Foraminifera, Evolutionary and Geological Events in the Context of Datum Planes. Pacific Neogene Datum Planes. University of Tokyo Press, Tokyo. 47–68 |
| Adams, C. G., Haak, R., 1962. The Stratigraphical Succession in the Batu Gading Area, Middle Baram, North Sarawak. In: The Geology and Mineral Resources of the Suai-Baram Area, North Sarawak. Geological Survey Department British Territories, Borneo. 141–150 |
| Adams, C. G., Frame, P., 1979. Observations on Cycloclypeus (Cycloclypeus) Carpenter and Cycloclypeus (Katacycloclypeus) Tan (Foraminifera). Bulletin of the British Museum of Natural History (Geology), 32(1): 3–17 |
| Allan, T., Trotter, J. A., Whitford, D. J., et al., 2000. Strontium Isotope Stratigraphy and the Oligocene-Miocene T-Letter "Stages" in Papua New Guinea. Papua New Guinea's Petroleum Industry in the 21st Century: Proceedings of the Fourth PNG Petroleum Convention. May 29–31, 2000, Port Moresby, PNG. 155–168 |
| Amato, V., Drooger, C. W., 1969. How to Measure the Angle γ in the Miogypsinidae. Revista Española de Micropaleontologia, 1(1): 19–244 |
| Banner, F. T., Hodgkinson, R. L., 1991. A Revision of the Foraminiferal Subfamily Heterostegininae. Revista Española de Micropaleontologia, XXIII(2): 101–140 |
| Banner, F. T., Samuel, M. A., 1995. Alanlordia, a New Genus of Acervuline Foraminifera from the Neogene of Indonesia. Journal of Micropalaeon-tology, 14(2): 107–117. https://doi.org/10.1144/jm.14.2.107 |
| Berggren, W. A., Kent, D. V., Flynn, J. J., 1985. Jurassic to Paleogene: Part 2. Palaeogene Geochronology and Chronostratigraphy. In: Snelling, N. J., ed., The Chronology of the Geological Record (Memoir 10). Geological Society of London, London. 141–186 |
| Berggren, W. A., Prothero, D. R., 1992. Eocene-Oligocene Climatic and Biotic Evolution: An Overview. In: Prothero, D. R., Berggren, W. A., eds., Eocene-Oligocene Climatic and Biotic Evolution. Princeton University Press, Princeton. 1–28 |
| Berggren, W. A., Kent, D. V., Swisher, C. C. III, et al., 1995. A Revised Cenozoic Geochronology and Chronostratigraphy. In: Berggren, W. A., Kent, D. V., Aubry, M. -P., et al., eds., Geochronology Time Scales and Global Correlation (54). SEPM Special Publication, 54: 129–212. https://doi.org/10.2110/pec.95.04.0129 |
| BouDagher-Fadel, M. K., Banner, F. T., 1999. Revision of the Stratigraphic Significance of the Oligocene-Miocene "Letter-Stages". Revue de Micropaléontologie, 42(2): 93–97. https://doi.org/10.1016/s0035-1598(99)90095-8 |
| BouDagher-Fadel, M. K., Price, G. D., 2013. The Phylogenetic and Palaeo-geographic Evolution of the Miogypsinid Larger Benthic Foraminifera. Journal of the Geological Society, 170(1): 185–208. https://doi.org/10.1144/jgs2011-149 |
| BouDagher-Fadel, M. K., Price, G. D., 2017. The Paleogeographic Evolution of the Orthophragminids of the Paleogene. Journal of Foraminiferal Research, 47(4): 337–357. https://doi.org/10.2113/gsjfr.47.4.337 |
| Bromfield, K., Renema, W., 2011. Comparison of 87Sr/86Sr Isotope and Biostratigraphic Ages of Uplifted Fossil Reefs in the Indo-Pacific: Indonesia, Papua New Guinea and Fiji. Australian Journal of Earth Sciences, 58(1): 61–73. https://doi.org/10.1080/08120099.2011.534816 |
| Cahuzac, B., Poignant, A., 1997. Essai de Biozonation de l'Oligo-Miocene dans les Bassins Européens à l'aide des Grands Foraminifères Néritiques. Bulletin de la Société Géologique de France, 168(2): 155–169 |
| Chaproniere, G. C. H., 1975. Palaeoecology of Oligo-Miocene Larger Foraminiferida, Australia. Alcheringa: An Australasian Journal of Pa-laeontology, 1(1): 37–58. https://doi.org/10.1080/03115517508619479 |
| Chaproniere, G. C. H., 1980. Biometrical Studies of Early Neogene Larger Foraminiferida from Australia and New Zealand. Alcheringa: An Australasian Journal of Palaeontology, 4(3): 153–181. https://doi.org/10.1080/03115518008618929 |
| Chaproniere, G. C. H., 1981. Australasian Mid-Tertiary Larger Foraminiferal Associations and Their Bearing on the East Indian Letter Classification. BMR Journal of Australian Geology & Geophysics, 6: 145–151 |
| Chaproniere, G. C. H., 1984. Oligocene and Miocene Larger Foraminiferida from Australia and New Zealand (Ⅵ). Australian Government Publishing Service, Canberra. 98 |
| Cole, W. S., 1954. Larger Foraminifera and Smaller Diagnostic Foraminifera from Bikini Drill Holes. U.S. Geological Survey Professional Paper, 260-O: 569–608 |
| Cole, W. S., 1957a. Larger Foraminifera from Eniwetok Drill Holes. U.S. Geological Survey Professional Paper, 260-V: 743–784 |
| Cole, W. S., 1957b. Larger foraminifera [of Saipan, Mariana Islands]. U.S. Geological Survey Professional Paper, 280-I: 321–357 |
| Cole, W. S., 1960. Upper Eocene and Oligocene Larger Foraminifera from Viti Levu, Fiji. U.S. Geological Survey Professional Paper, 374-A: 1–6 |
| Cole, W. S., 1963. Tertiary Larger Foraminifera from Guam. U.S. Geological Survey Professional Paper, 403-E: 1–28 |
| Cotton, L. J., Pearson, P. N., 2011. Extinction of Larger Benthic Foraminifera at the Eocene/Oligocene Boundary. Palaeogeography, Palaeoclimatology, Palaeoecology, 311(3/4): 281–296. https://doi.org/10.1016/j.palaeo.2011.09.008 |
| Cotton, L. J., Pearson, P. N., Renema, W., 2014. Stable Isotope Stratigraphy and Larger Benthic Foraminiferal Extinctions in the Melinau Limestone, Sarawak. Journal of Asian Earth Sciences, 79: 65–71. https://doi.org/10.1016/j.jseaes.2013.09.025 |
| Crotty, K. J., Engelhardt, D. W., 1993. Larger Foraminifera and Palyno-morphs of the Upper Malawa and Lower Tonasa Formations, South-western Sulawesi Island, Indonesia. Proceedings of the International Symposium on Biostratigraphy of Mainland Southeast Asia: Facies and Paleontology. Indonesian Petroleum Association. 71–82 |
| Drooger, C. W., 1963. Evolutionary Trends in Miogypsinidae. In: Evolutionary Trends in Foraminifera. Elsevier, Amsterdam. 315–349 |
| Drooger, C. W., 1993. Radial Foraminifera: Morphometrics and Evolution (41). Afd. Natuurkunde, Eerste Reeks, Deel 41, Amsterdam. 241 |
| Drooger, C. W., Raju, D. S. N., 1978. Early Miogypsinoides in Kutch, Western India (Ⅰ-Ⅱ). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen (B), 81: 186–219 |
| Fermont, W. J. J., 1977. Biometrical Investigations of the Genus Operculina in Recent Sediments of the Gulf of Eliat. Utrecht Micropaleontological Bulletins 15: 149–164 |
| Fermont, W. J. J., 1982. Discocydinidae from Ein Avedat (Israel). Utrecht Micropaleontological Bulletins, 27: 1–173 |
| Gold, D. P., White, L. T., Gunawan, I., et al., 2017. Relative Sea-Level Change in Western New Guinea Recorded by Regional Biostratigraphic Data. Marine and Petroleum Geology, 86: 1133–1158. https://doi.org/10.1016/j.marpetgeo.2017.07.016 |
| Gould, S. J., Eldredge, N., 1977. Punctuated Equilibria: The Tempo and Mode of Evolution Reconsidered. Paleobiology, 3(2): 115–151. https://doi.org/10.1017/s0094837300005224 |
| Gradstein, F. M., Ogg, J. G., Smith, A. G., 2004. A Geologic Time Scale 2004. Cambridge University Press, Cambridge. 500 |
| Hassl, D. A., Hansen, T. A., 1996. Timing of Latest Eocene Molluscan Extinction Patterns in Mississppi. PALAIOS, 11(5): 487–494. https://doi.org/10.2307/3515214 |
| Hallock, P., 1985. Why are Larger Foraminifera Large? Paleobiology, 11(2): 195–208. https://doi.org/10.1017/s0094837300011507 |
| Hallock, P., Glenn, E. C., 1986. Larger Foraminifera: A Tool for Paleoenvironmental Analysis of Cenozoic Carbonate Depositional Facies. PALAIOS, 1(1): 55. https://doi.org/10.2307/3514459 |
| Hallock, P., Premoli Silva, I., Boersma, A., 1991. Similarities between Planktonic and Larger Foraminiferal Evolutionary Trends through Paleogene Paleoceanographic Changes. Palaeogeography, Palaeoclima-tology, Palaeoecology, 83(1/2/3): 49–64. https://doi.org/10.1016/0031-0182(91)90075-3 |
| Hansen, T. A., 1987. Extinction of Late Eocene to Oligocene Molluscs: Relationship to Shelf Area, Temperature Changes, and Impact Events. PALAIOS, 2(1): 69. https://doi.org/10.2307/3514573 |
| Haq, B. U., Al-Qahtani, A. M., 2005. Phanerozoic Cycles of Sea-Level Change on the Arabian Platform. GeoArabia 10: 127–160 doi: 10.2113/geoarabia1002127 |
| Hashimoto, W., Matsumaru, K., Kurihara, K., 1977. Larger Foraminifera from the Philippines, V. Larger Foraminifera from Cenozoic Limestones in the Mansalay Vicinity, Oriental Mindoro, with an Appendix "An Orbitoid-Bearing Limestone from Barahid, Bongabong. Geology and Palaeontology of Southeast Asia, 18: 59–76 |
| Hohenegger, J., 2000. Remarks on West Pacific Nummulitidae (Foraminifera). The Journal of Foraminiferal Research, 30(1): 3–28. https://doi.org/10.2113/0300003 |
| Ho, K. F., 1973. Morphometric Trend in Lepidocyclina and Its Application to Time Stratigraphy in N.W. Borneo. Shell Report Exp. 529 |
| Hottinger, L., 1977. Foraminiferes Operculiniformes. Memoires du Museum National d'Histoire Naturelle, Ser. C, Sei. Terre, 40: 1–159 |
| Hottinger, L., Romero, J., Caus, E., 2001. Architecture and Revision of the Pellatispirines, Planispiral Canaliferous Foraminifera from the Late Eocene Tethys. Micropaleontology, 47(2): 35–77 |
| Krijnen, W. F., 1931. Het Genus Spiroclypeus in her Indo-Pacifische Gebied. Verhandelingen van het Geologisch en Mijnbouwkundig Genootschap voor Nederland en Koloniën, Ⅺ: 77–112 |
| Laagland, H., 1990. Cycloclypeus in the Mediterranean Oligocene. Utrecht Micropaleontological Bulletins, 39: 171 |
| Lapre, J. F., Thornton, M. S., 1970. Carbonate Stratigraphy of the SE Platform Complex, Central Luconia Province. Shell report Exp. R. 1741 |
| Less, G., Özcan, E., 2008. The Late Eocene Evolution of Nummulitid Foraminifer Spiroclypeus in the Western Tethys. Acta Palaeontologica Polonica, 53(2): 303–316. https://doi.org/10.4202/app.2008.0211 |
| Leupold, W., van der Vlerk, I. M., 1931. The Tertiary. Leidsche Geologische Mededelingen, 5: 611–648 |
| Levit, G. S., Olsson, L., 2006. "Evolution on Rails": Mechanisms and Levels of Orthogenesis. Annals of the History and Philosophy of Biology, 11: 99–138 |
| Liechti, P., 1960. The Geology of Sarawak, Brunei and the Western Part of North Borneo (Bulletin 3). Geological Survey Department British Territories in Borneo, Kuching Sarawak |
| Lunt, P., 1984. The Biostratigraphy and Palaeoecology of the Oligo-Miocene Terra Limestone, Western Cyprus: [Dissertation]. University College London, London. 90 |
| Lunt, P., 2003. Biogeography of some Eocene Larger Foraminifera, and Their Application in Distinguishing Geological Plates. Palaeontologica Electronica, 6(1): 1–22 |
| Lunt, P., 2013. The Sedimentary Geology of Java. Indonesian Petroleum Association, Jakarta. 346 |
| Lunt, P., 2021a. Biological Evolution of SE Asian Carbonates, Based on Their Microfossil Content. SEPM Special Publication, Cenozoic Isolated Carbonate Platforms-Focus Southeast Asia, https://doi.org/10.2110/sepmsp.114.07 |
| Lunt, P., 2021b. Tectono-Stratigraphic Framework of Luconia Carbonates. SEPM Special Publication, Cenozoic Isolated Carbonate Platforms-Focus Southeast Asia, https://doi.org/10.2110/sepmsp.114.08 |
| Lunt, P., Allan, T., 2004. A History and Application of Larger Foraminifera in Indonesian Biostratigraphy, Calibrated to Isotopic Dating. GRDC Workshop on Micropalaeontology. 112 |
| Lunt, P., Burgon, G., Baky, A., 2009. The Pemali Formation of Central Java and Equivalents: Indicators of Sedimentation on an Active Plate Margin. Journal of Asian Earth Sciences, 34(1): 100–113. https://doi.org/10.1016/j.jseaes.2008.03.006 |
| Lunt, P., Renema, W., 2014. On the Heterostegina-Tansinhokella-Spiroclypeus Lineages in SE Asia. Berita Sedimentologi, 30: 6–31 |
| Lunt, P., Woodroof, P., 2021. Tectono-Stratigraphic Controls on Cenozoic SE Asian Carbonates. SEPM Special Publication, Cenozoic Isolated Carbonate Platforms-Focus Southeast Asia, https://doi.org/10.2110/sepmsp.114.06 |
| Lyell, C., 1833. Principles of Geology, Volumes 1–3. John Murray, London |
| MacArthur, R., Wilson, E. O., 1967. The Theory of Island Biogeography. Princeton University Press |
| McArthur, J. M., Howarth, R. J., 2004. Strontium Isotope Stratigraphy. In: Gradstein, F. M., Ogg, J. G., Smith, A. G., eds., A Geologic Time Scale 2004. Cambridge University Press, Cambridge. 96–105 |
| Mohler, W. A., 1950. Flosculinella reicheli n. sp. aus dem Tertiär e5 von Borneo. Schweiz. Naturf. Ges., Verh. (Soc. Helv. Sci. Nat. Actes). 129 |
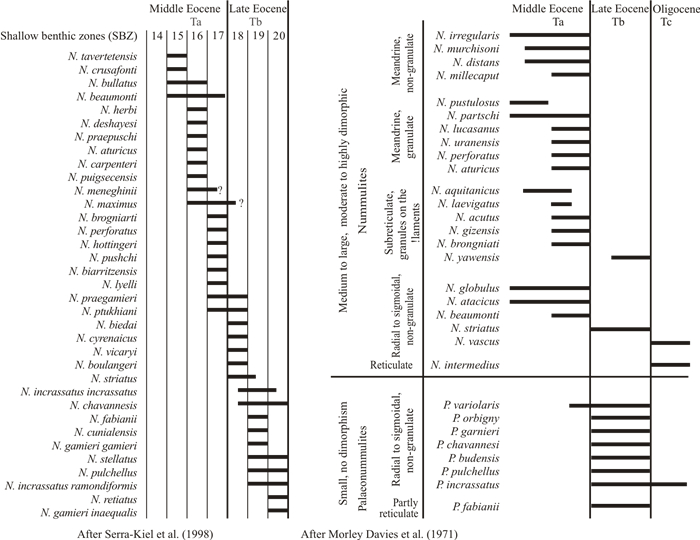
| Morley Davies, A., Eames, F. E., Savage, R. J. G., 1971. Tertiary Faunas. George Allen & Unwin Ltd., London. 1018 |
| Morley, R. J., 2000. Origin and Evolution of Tropical Rain Forests. Wiley & Sons, London. 362 |
| Moss, S. J., Chambers, J. L. C., 1999. Tertiary Facies Architecture in the Kutai Basin, Kalimantan, Indonesia. Journal of Asian Earth Sciences, 17(1/2): 157–181. https://doi.org/10.1016/s0743-9547(98)00035-x |
| Muhar, A., 1957. Micropalaeontologishe Onderzoek van Monsters Afkomstig van het Geologishe Onderzoek Tuban. B.P.M. Unpublished Report Gr. Sb. 1770: 22 |
| Ozawa, T., 1975. Evolution of Lepidolina Multiseptata (Permian Foraminifer) in East Asia. Memoirs of the Faculty of Science, Kyūsyū University Series D, Geology, 23(2): 117–164. https://doi.org/10.5109/1544173 |
| Özcan, E., Less, G., 2009. First Record of the Co-Occurrence of Western Tethyan and Indo-Pacific Larger Foraminifera in the Burdigalian of the Mediterranean Province. The Journal of Foraminiferal Research, 39(1): 23–39. https://doi.org/10.2113/gsjfr.39.1.23 |
| Özcan, E., Less, G., Báldi-Beke, M., et al., 2009. Oligo-Miocene Foraminiferal Record (Miogypsinidae, Lepidocyclinidae and Nummu-litidae) from the Western Taurides (SW Turkey): Biometry and Implications for the Regional Geology. Journal of Asian Earth Sciences, 34(6): 740–760. https://doi.org/10.1016/j.jseaes.2008.11.002 |
| Perrin, C., 2002. Tertiary: The Emergence of Modern Reef Ecosystems. In: Kiessling, W., Flügel, E., Golonka, J., eds., Phanerozoic Reef Patterns. SEPM Special Publication, 72: 587–621 |
| Perch-Nielsen, K., 1985. Cenozoic Calcareous Nannofossils. In: Bolli, H. M., Saunders, J. B., Perch-Nielsen, K., eds., Plankton Stratigraphy. Cambridge University Press, Cambridge. 427–554 |
| Prothero, D. R., 1994. The Late Eocene-Oligocene Extinctions. Annual Review of Earth and Planetary Sciences, 22: 145–165. https://doi.org/10.1146/annurev.ea.22.050194.001045 |
| Prothero, D. R., Swisher, C. C., 1992. Magnetostratigraphy and Geochrono-logy of the Terrestrial Eocene-Oligocene Transition in North America. Eocene-Oligocene Climatic and Biotic Evolution. Princeton University Press. 46–73. https://doi.org/10.1515/9781400862924.46 |
| Racey, A., 1994. Biostratigraphy and Palaeobiogeographic Significance of Tertiary Nummulitids (Foraminifera) from Northern Oman. In: Simmons, M. D., ed., Micropalaeontology and Hydrocarbon Exploation in the Middle East. Chapman and Hall, London. 343–367 |
| Racey, A., 1995. Lithostratigraphy and Larger Foraminiferal (Nummulitid) Biostratigraphy of the Tertiary of Northern Oman. Micropaleontology, 41: 1–123. https://doi.org/10.2307/1485849 |
| Raju, D. S. N., 1974. Study of Indian Miogypsinidae. Utrecht Micropaleontological Bulletins, 9: 148 |
| Renema, W., 2002. Larger Foraminfera as Marine Environmental Indicators. Scripta Geologica, 124: 1–260 |
| Renema, W., 2015. Spatiotemporal Variation in Morphological Evolution in the Oligocene-Recent Larger Benthic Foraminifera Genus Cycloclypeus Reveals Geographically Undersampled Speciation. GeoResJ, 5: 12–22. https://doi.org/10.1016/j.grj.2014.11.001 |
| Renema, W., Troelstra, S. R., 2001. Larger Foraminifera Distribution on a Mesotrophic Carbonate Shelf in SW Sulawesi (Indonesia). Palaeogeography, Palaeoclimatology, Palaeoecology, 175(1/2/3/4): 125–146. https://doi.org/10.1016/s0031-0182(01)00389-3 |
| Renema, W., Bellwood, D. R., Braga, J. C., et al., 2008. Hopping Hotspots: Global Shifts in Marine Biodiversity. Science, 321(5889): 654–657. https://doi.org/10.1126/science.1155674 |
| Renema, W., Warter, V., Novak, V., et al., 2015. Ages of Miocene Fossil Localities in the Northern Kutai Basin (East Kalimantan, Indonesia). PALAIOS, 30(1): 26–39. https://doi.org/10.2110/palo.2013.127 |
| Sarkar, S., 2018. The Enigmatic Palaeocene-Eocene Coralline Distichoplax: Approaching the Structural Complexities, Ecological Affinities and Extinction Hypotheses. Marine Micropaleontology, 139: 72–83. https://doi.org/10.1016/j.marmicro.2017.12.001 |
| Saw, B. B., Schlaich, M., Pöppelreiter, M. C., et al., 2019. Facies, Depositional Environments, and Anatomy of the Subis Build-up in Sarawak, Malaysia: Implications on other Miocene Isolated Carbonate Build-ups. Facies, 65(3): 1–14. https://doi.org/10.1007/s10347-019-0571-6 |
| Schaub, H., 1981. Nummulites et Assilines de la Tethys Paleogene: Taxinomie, Phylogenese et Biostratigraphie. Memoires Suisses de Paleontologie, 104/105/106: 1–236 |
| Serra-Kiel, J., Hottinger, L., Caus, E., et al., 1998. Larger Foraminiferal Biostratigraphy of the Tethyan Paleocene and Eocene. Bulletin de la Societe Geologique de France, 169(2): 281–299 |
| Serra-Kiel, J., Gallardo-Garcia, A., Razin, P., et al., 2016. Middle Eocene-Early Miocene Larger Foraminifera from Dhofar (Oman) and Socotra Island (Yemen). Arabian Journal of Geosciences, 9(5): 1–95. https://doi.org/10.1007/s12517-015-2243-3 |
| Singh, S. D., Raju, D. S. N., 2018. Morphometric Data on Miogypsina (Lepidosemicyclina) Bifida, Foraminifera from L-Ⅲ Reservoir, Mumbai Offshore, India. Journal of the Geological Society of India, 91(3): 329–333. https://doi.org/10.1007/s12594-018-0858-1 |
| Stanley, S. M., Hardie, L. A., 1998. Secular Oscillations in the Carbonate Mineralogy of Reef-Building and Sediment-Producing Organisms Driven by Tectonically Forced Shifts in Seawater Chemistry. Palaeogeography, Palaeoclimatology, Palaeoecology, 144(1/2): 3–19. https://doi.org/10.1016/s0031-0182(98)00109-6 |
| Stearns, S. C., 1992. The Evolution of Life Histories. Oxford University Press, Oxford |
| Swisher, C. C. III, Prothero, D. R., 1990. Single-Crystal 40Ar/39Ar Dating of the Eocene-Oligocene Transition in North America. Science, 249(4970): 760–762. https://doi.org/10.1126/science.249.4970.760 |
| Tan, S. H., 1932. On the Genus Cycloclypeus Carpenter Part 1. and an Appendix on the Heterostegines of Tjimanggoe, S. Bantam, Java. Wetenschappelijke Mededelingen Dienst van den Mijnbouw in Nederlandsch-Indië, 19: 1–194 |
| Vahrenkamp, V. C., 1996. Growth and Demise of the Miocene Central Luconia Carbonate Province: Implications for Regional Geology and Reservoir Production Behaviour. Sarawak Shell Report |
| van Bemmelen, R. W., 1949. The Geology of Indonesia. Government Printing Office, Hague, Netherlands. 997 |
| van der Vlerk, I. M., 1924. Foraminiferen uit het Tertiair van Java. Weteenschappelijke Mededeelingen Dienst Mijnbouw Ned. Indie, 1: 1–13 |
| van der Vlerk, I. M., 1959. Problems and Principles of Tertiary and Quaternary Stratigraphy. Quarterly Journal of the Geological Society, 115(1/2/3/4): 49–64. https://doi.org/10.1144/gsl.jgs.1959.115.01.04 |
| van der Vlerk, I. M., 1963. Biometric Research on Lepidocyclina. Micropaleontology, 9: 425–426. https://doi.org/10.2307/1484502 |
| van der Vlerk, I. M., 1973. An Improved Method of Biometric Research. Proc. Kon. Ned. Akad. Wet., B76(4): 245–259 |
| van der Vlerk, I. M., Umbgrove, J. H. F., 1927. Tertiaire Gidsforaminiferen uit Nederlandsch Oost-Indië. Wetenschappelijke Mededelingen Dienst van den Mijnbouw in Nederlandsch-Indië, 6: 1–31 |
| van der Vlerk, I. M., Postuma, J. A., 1967. Oligo-Miocene Lepidocyclinas and Planktonic Foraminifera from East Java and Madura. Proceedings of the Koninklijke Nederlandse Academie van Wetenschappen, 70(B): 392–399 |
| van Gorsel, J. T., Lunt, P., Morley, R. J., 2014. Introduction to Cenozoic Biostratigraphy of Indonesia-SE Asia. Berita Sedimentologi, 29: 4–40 |
| van Vessem, E. J., 1978. Study of Lepidocylinidae from South East Asia, particularly from Java and Borneo. Utrecht Micropaleontological Bulletins, 19: 1–163 |
| Visser, W. A., Hermes, J. J., 1962. Geological Results of the Search for Oil in Netherlands New Guinea (20). Nederlandsche Nieuw Guinee Petroleum Maatschappij, Deltf |
| Wade, B. S., 2004. Planktonic Foraminiferal Biostratigraphy and Mechanisms in the Extinction of Morozovella in the Late Middle Eocene. Marine Micropaleontology, 51(1/2): 23–38. https://doi.org/10.1016/j.marmicro.2003.09.001 |
| Wade, B. S., Pearson, P. N., Berggren, W. A., et al., 2011. Review and Revision of Cenozoic Tropical Planktonic Foraminiferal Biostrati-graphy and Calibration to the Geomagnetic Polarity and Astronomical Time Scale. Earth-Science Reviews, 104(1/2/3): 111–142. https://doi.org/10.1016/j.earscirev.2010.09.003 |
| Westerhold, T., Marwan, N., Drury, A. J., et al., 2020. An Astronomically Dated Record of Earth's Climate and Its Predictability over the Last 66 Million Years. Science, 369(6509): 1383–1387. https://doi.org/10.1126/science.aba6853 |
| White, L. T., Hall, R., Armstrong, R. A., et al., 2017. The Geological History of the Latimojong Region of Western Sulawesi, Indonesia. Journal of Asian Earth Sciences, 138: 72–91. https://doi.org/10.1016/j.jseaes.2017.02.005 |
| Wilson, M. E. J., 2002. Cenozoic Carbonates in Southeast Asia: Implications for Equatorial Carbonate Development. Sedimentary Geology, 147(3/4): 295–428. https://doi.org/10.1016/S0037-0738(01)00228-7 |
| Wilson, M. E. J., 2008. Global and Regional Influences on Equatorial Shallow-Marine Carbonates during the Cenozoic. Palaeogeography, Palaeoclimatology, Palaeoecology, 265(3/4): 262–274. https://doi.org/10.1016/j.palaeo.2008.05.012 |
| Yabe, H., Hanzawa, S., 1928. Tertiary Foraminiferous Rocks of Taiwan (Formosa). Proceedings of the Imperial Academy, 4(9): 533–536. https://doi.org/10.2183/pjab1912.4.533 |
| Zeiza, A., Simaeys, V. S., Musgrove, F., et al., 2012. The Impact of Differential Subsidence Rates in Shallow Water Carbonate, Reservoir Quality: An Example from the East Java Basin, Indonesia. Proceedings Indonesian Petroleum Association Convention, 36: 1–13 |
Figures(6) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: