| Citation: | Mingxian Han, Jianrong Huang, Jian Yang, Beichen Wang, Sun Xiaoxi, Hongchen Jiang. Distinct Assembly Mechanisms for Prokaryotic and Microeukaryotic Communities in the Water of Qinghai Lake. Journal of Earth Science, 2023, 34(4): 1189-1200. doi: 10.1007/s12583-023-1812-8

|
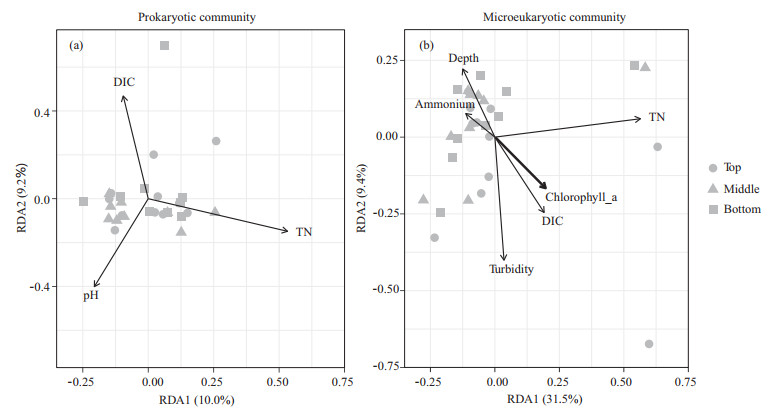
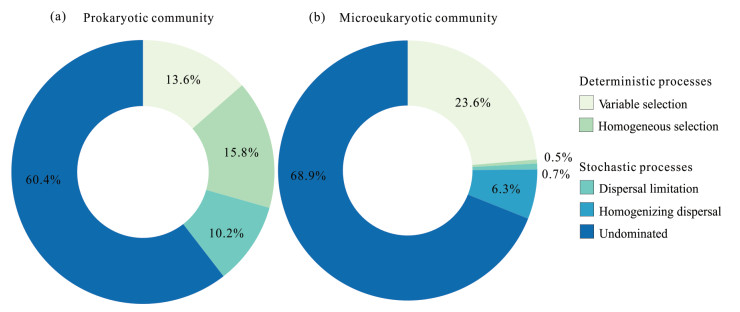
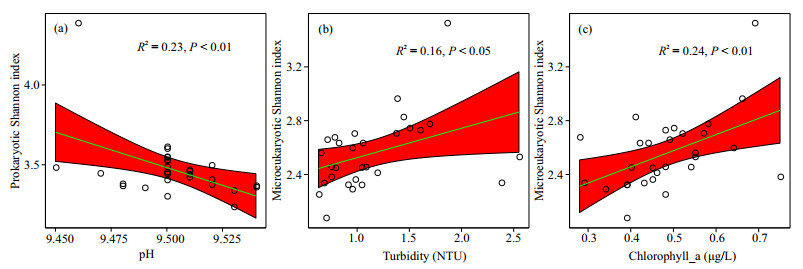
Assembly processes of prokaryotic and microeukaryotic community is an important issue in microbial ecology. However, unclear remains about the relative contribution of deterministic and stochastic processes to the shaping of prokaryotic and microeukaryotic communities in saline lake water. Here, we systematically investigated the assembly processes governing the prokaryotic and microeukaryotic communities in Qinghai Lake with the use of Illumina sequencing and a null model. The results showed that both deterministic and stochastic processes play vital roles in shaping the assemblies of prokaryotic and microeukaryotic communities, in which stochastic processes appeared to dominate (> 70%). Prokaryotic communities were mainly governed by non-dominant processes (60.4%), followed by homogeneous selection (15.8%), variable selection (13.6%) and dispersal limitation (10.2%), whereas microeukaryotes were strongly driven by non-dominant processes (68.9%), followed by variable selection (23.6%) and homogenizing dispersal (6.3%). In terms of variable selection, nutrients (e.g., ammonium, dissolved inorganic carbon, dissolved organic carbon and total nitrogen) were the major factors influencing prokaryotic and microeukaryotic community structures. In summary, prokaryotes and microeukaryotes can be predominantly structured by different assembly mechanisms, in which stochasticity is stronger than deterministic processes. This finding helps to better comprehend the assembly of prokaryotic and eukaryotic communities in saline lakes.

| Bertolet, B. L., Louden, S. I., Jones, S. E., 2022. Microbial Community Composition, and not pH, Influences Lake Sediment Function. Ecosphere, 13(5): e4091. https://doi.org/10.1002/ecs2.4091 |
| Bolyen, E., Rideout, J. R., Dillon, M. R., et al., 2019. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nature Biotechnology, 37(8): 852–857. https://doi.org/10.1038/s41587-019-0209-9 |
| Bradshaw, A. L., Brewer, P. G., Shafer, D. K., et al., 1981. Measurements of Total Carbon Dioxide and Alkalinity by Potentiometric Titration in the GEOSECS Program. Earth and Planetary Science Letters, 55(1): 99–115. https://doi.org/10.1016/0012-821x(81)90090-x |
| Callahan, B. J., McMurdie, P. J., Holmes, S. P., 2017. Exact Sequence Variants should Replace Operational Taxonomic Units in Marker-Gene Data Analysis. The ISME Journal, 11(12): 2639–2643. https://doi.org/10.1038/ismej.2017.119 |
| Caporaso, J. G., Lauber, C. L., Walters, W. A., et al., 2012. Ultra-High-Throughput Microbial Community Analysis on the Illumina HiSeq and MiSeq Platforms. The ISME Journal, 6(8): 1621–1624. https://doi.org/10.1038/ismej.2012.8 |
| Caruso, T., Chan, Y., Lacap, D. C., et al., 2011. Stochastic and Deterministic Processes Interact in the Assembly of Desert Microbial Communities on a Global Scale. The ISME Journal, 5(9): 1406–1413. https://doi.org/10.1038/ismej.2011.21 |
| Chase, J. M., 2010. Stochastic Community Assembly Causes Higher Biodiversity in More Productive Environments. Science, 328(5984): 1388–1391. https://doi.org/10.1126/science.1187820 |
| Chase, J. M., Myers, J. A., 2011. Disentangling the Importance of Ecological Niches from Stochastic Processes across Scales. Philosophical Transactions of the Royal Society B: Biological Sciences, 366(1576): 2351–2363. https://doi.org/10.1098/rstb.2011.0063 |
| Chen, W. D., Ren, K. X., Isabwe, A., et al., 2019. Stochastic Processes Shape Microeukaryotic Community Assembly in a Subtropical River across Wet and Dry Seasons. Microbiome, 7(1): 138. https://doi.org/10.1186/s40168-019-0749-8 |
| Cole, J. J., Findlay, S., Pace, M. L., 1988. Bacterial Production in Fresh and Saltwater Ecosystems: A Cross-System Overview. Marine Ecology Progress Series, 43: 1–10. https://doi.org/10.3354/meps043001 |
| Cui, B. L., Li, X. Y., 2016. The Impact of Climate Changes on Water Level of Qinghai Lake in China over the Past 50 Years. Hydrology Research, 47(2): 532–542. https://doi.org/10.2166/nh.2015.237 |
| Danger, M., Gessner, M. O., Bärlocher, F., 2016. Ecological Stoichiometry of Aquatic Fungi: Current Knowledge and Perspectives. Fungal Ecology, 19: 100–111. https://doi.org/10.1016/j.funeco.2015.09.004 |
| Demoling, F., Nilsson, L. O., Bååth, E., 2008. Bacterial and Fungal Response to Nitrogen Fertilization in Three Coniferous Forest Soils. Soil Biology and Biochemistry, 40(2): 370–379. https://doi.org/10.1016/j.soilbio.2007.08.019 |
| Dini-Andreote, F., Stegen, J. C., van Elsas, J. D., et al., 2015. Disentangling Mechanisms that Mediate the Balance between Stochastic and Deterministic Processes in Microbial Succession. Proceedings of the National Academy of Sciences of the United States of America, 112(11): E1326–E1332. https://doi.org/10.1073/pnas.1414261112 |
| Dong, H. L., Zhang, G. X., Jiang, H. C., et al., 2006. Microbial Diversity in Sediments of Saline Qinghai Lake, China: Linking Geochemical Controls to Microbial Ecology. Microbial Ecology, 51(1): 65–82. https://doi.org/10.1007/s00248-005-0228-6 |
| Fabian, J., Zlatanovic, S., Mutz, M., et al., 2017. Fungal-Bacterial Dynamics and Their Contribution to Terrigenous Carbon Turnover in Relation to Organic Matter Quality. The ISME Journal, 11(2): 415–425. https://doi.org/10.1038/ismej.2016.131 |
| Farjalla, V. F., Srivastava, D. S., Marino, N. A. C., et al., 2012. Ecological Determinism Increases with Organism Size. Ecology, 93(7): 1752–1759. https://doi.org/10.1890/11-1144.1 |
| Finlay, B. J., 2002. Global Dispersal of Free-Living Microbial Eukaryote Species. Science, 296(5570): 1061–1063. https://doi.org/10.1126/science.1070710 |
| Frade, P. R., Glasl, B., Matthews, S. A., et al., 2020. Spatial Patterns of Microbial Communities across Surface Waters of the Great Barrier Reef. Communications Biology, 3(1): 442. https://doi.org/10.1038/s42003-020-01166-y |
| Gad, M., Hou, L. Y., Li, J. W., et al., 2020. Distinct Mechanisms Underlying the Assembly of Microeukaryotic Generalists and Specialists in an Anthropogenically Impacted River. Science of the Total Environment, 748: 141434. https://doi.org/10.1016/j.scitotenv.2020.141434 |
| Ge, Y. H., Meng, X. L., Heino, J., et al., 2021. Stochasticity Overrides Deterministic Processes in Structuring Macroinvertebrate Communities in a Plateau Aquatic System. Ecosphere, 12(7): e03675. https://doi.org/10.1002/ecs2.3675 |
| Gu, Z. Q., Liu, K. S., Pedersen, M. W., et al., 2021. Community Assembly Processes Underlying the Temporal Dynamics of Glacial Stream and Lake Bacterial Communities. Science of the Total Environment, 761: 143178. https://doi.org/10.1016/j.scitotenv.2020.143178 |
| Hammer, U. T., 1981. 5. Primary Production in Saline Lakes. Hydrobiologia, 81(1): 47–57. https://doi.org/10.1007/BF00048705 |
| Hanashiro, F. T. T., De Meester, L., Vanhamel, M., et al., 2022. Bacterioplankton Assembly along a Eutrophication Gradient is Mainly Structured by Environmental Filtering, Including Indirect Effects of Phytoplankton Composition. Microbial Ecology, 1–11. https://doi.org/10.1007/s00248-022-01994-x |
| Hubbell, S. P., 2011. The Unified Neutral Theory of Biodiversity and Biogeography (MPB-32). Princeton University Press, Princeton. 32 |
| Hussain, S., Liu, H., Liu, S. L., et al., 2021. Distribution and Assembly Processes of Soil Fungal Communities along an Altitudinal Gradient in Tibetan Plateau. Journal of Fungi (Basel, Switzerland), 7(12): 1082. https://doi.org/10.3390/jof7121082 |
|
Jellison, R., Williams, W. D., Timms, B., et al., 2008. Salt Lakes: Values, Threats and Future. In: Polunin, N. V. C., ed., Aquatic Ecosystems: Trends and Global Prospects. Cambridge University Press, Cambridge. 94–110. |
| Jiang, H. C., Dong, H. L., Yu, B. S., et al., 2008. Dominance of Putative Marine Benthic Archaea in Qinghai Lake, North-Western China. Environmental Microbiology, 10(9): 2355–2367. https://doi.org/10.1111/j.1462-2920.2008.01661.x |
| Jiao, S., Yang, Y. F., Xu, Y. Q., et al., 2020. Balance between Community Assembly Processes Mediates Species Coexistence in Agricultural Soil Microbiomes across Eastern China. The ISME Journal, 14(1): 202–216. https://doi.org/10.1038/s41396-019-0522-9 |
| Koorem, K., Gazol, A., Öpik, M., et al., 2014. Soil Nutrient Content Influences the Abundance of Soil Microbes but not Plant Biomass at the Small-Scale. PLoS One, 9(3): e91998. https://doi.org/10.1371/journal.pone.0091998 |
| Kuznetsov, S. I., 1975. The Microflora of Lakes and Its Geochemical Activity. University of Texas Press, Austin. https://doi.org/10.7560/750104 |
| López-García, P., Rodríguez-Valera, F., Pedrós-Alió, C., et al., 2001. Unexpected Diversity of Small Eukaryotes in Deep-Sea Antarctic Plankton. Nature, 409(6820): 603–607. https://doi.org/10.1038/35054537 |
| Larsen, P. E., Field, D., Gilbert, J. A., 2012. Predicting Bacterial Community Assemblages Using an Artificial Neural Network Approach. Nature Methods, 9(6): 621–625. https://doi.org/10.1038/nmeth.1975 |
| Li, X. Y., Shi, F. Z., Ma, Y. J., et al., 2022. Significant Winter CO2 Uptake by Saline Lakes on the Qinghai-Tibet Plateau. Global Change Biology, 28(6): 2041–2052. https://doi.org/10.1111/gcb.16054 |
| Liu, K. S., Liu, Y. Q., Hu, A. Y., et al., 2020. Different Community Assembly Mechanisms Underlie Similar Biogeography of Bacteria and Microeukaryotes in Tibetan Lakes. FEMS Microbiology Ecology, 96(6): fiaa071. https://doi.org/10.1093/femsec/fiaa071 |
| Liu, L. M., Yang, J., Zhang, Y. Y., 2011. Genetic Diversity Patterns of Microbial Communities in a Subtropical Riverine Ecosystem (Jiulong River, Southeast China). Hydrobiologia, 678(1): 113–125. https://doi.org/10.1007/s10750-011-0834-x |
| Liu, M. H., Sui, X., Hu, Y. B., et al., 2019. Microbial Community Structure and the Relationship with Soil Carbon and Nitrogen in an Original Korean Pine Forest of Changbai Mountain, China. BMC Microbiology, 19(1): 218. https://doi.org/10.1186/s12866-019-1584-6 |
| Liu, Y. Q., Yao, T. D., Jiao, N. Z., et al., 2013. Salinity Impact on Bacterial Community Composition in Five High-Altitude Lakes from the Tibetan Plateau, Western China. Geomicrobiology Journal, 30(5): 462–469. https://doi.org/10.1080/01490451.2012.710709 |
| Logares, R., Tesson, S. V. M., Canbäck, B., et al., 2018. Contrasting Prevalence of Selection and Drift in the Community Structuring of Bacteria and Microbial Eukaryotes. Environmental Microbiology, 20(6): 2231–2240. https://doi.org/10.1111/1462-2920.14265 |
| Ma, R. H., Yang, G. S., Duan, H. T., et al., 2011. China's Lakes at Present: Number, Area and Spatial Distribution. Science China Earth Sciences, 54(2): 283–289. https://doi.org/10.1007/s11430-010-4052-6 |
| Ma, S. C., Fang, J. H., Liu, J., et al., 2022. Differences in Sediment Carbon-Fixation Rate and Associated Bacterial Communities in Four Wetland Types in Hulun Lake Basin. CATENA, 213: 106167. https://doi.org/10.1016/j.catena.2022.106167 |
| Ma, X. J., Chen, T., Zhang, G. S., et al., 2004. Microbial Community Structure along an Altitude Gradient in Three Different Localities. Folia Microbiologica, 49(2): 105–111. https://doi.org/10.1007/BF02931382 |
| Ma, Y. T., Li, J. Q., Wu, J., et al., 2018. Bacterial and Fungal Community Composition and Functional Activity Associated with Lake Wetland Water Level Gradients. Scientific Reports, 8(1): 760. https://doi.org/10.1038/s41598-018-19153-z |
| Martiny, J. B. H., Bohannan, B. J. M., Brown, J. H., et al., 2006. Microbial Biogeography: Putting Microorganisms on the Map. Nature Reviews Microbiology, 4(2): 102–112. https://doi.org/10.1038/nrmicro1341 |
| Massana, R., Logares, R., 2013. Eukaryotic Versus Prokaryotic Marine Picoplankton Ecology. Environmental Microbiology, 15(5): 1254–1261. https://doi.org/10.1111/1462-2920.12043 |
| Monchamp, M. E., Spaak, P., Pomati, F., 2019. High Dispersal Levels and Lake Warming Are Emergent Drivers of Cyanobacterial Community Assembly in Peri-Alpine Lakes. Scientific Reports, 9(1): 7366. https://doi.org/10.1038/s41598-019-43814-2 |
| Neal, C., Neal, M., Wickham, H., 2000. Phosphate Measurement in Natural Waters: Two Examples of Analytical Problems Associated with Silica Interference Using Phosphomolybdic Acid Methodologies. Science of the Total Environment, 251/252: 511–522. https://doi.org/10.1016/S0048-9697(00)00402-2 |
| Ning, D. L., Yuan, M. T., Wu, L. W., et al., 2020. A Quantitative Framework Reveals Ecological Drivers of Grassland Microbial Community Assembly in Response to Warming. Nature Communications, 11(1): 4717. https://doi.org/10.1038/s41467-020-18560-z |
| Oksanen, J., Blanchet, F. G., Friendly, M., et al., 2020. Vegan: Community Ecology Package, R Package Version 2.5–7 |
| Oren, A., 2008. Microbial Life at High Salt Concentrations: Phylogenetic and Metabolic Diversity. Saline Systems, 4: 2. https://doi.org/10.1186/1746-1448-4-2 |
| Paul Antony, C., Kumaresan, D., Hunger, S., et al., 2013. Microbiology of Lonar Lake and other Soda Lakes. The ISME Journal, 7(3): 468–476. https://doi.org/10.1038/ismej.2012.137 |
| Powell, J. R., Karunaratne, S., Campbell, C. D., et al., 2015. Deterministic Processes Vary During Community Assembly for Ecologically Dissimilar Taxa. Nature Communications, 6(1): 8444. https://doi.org/10.1038/ncomms9444 |
| Ramoneda, J., Le Roux, J. J., Frossard, E., et al., 2020. Experimental Assembly Reveals Ecological Drift as a Major Driver of Root Nodule Bacterial Diversity in a Woody Legume Crop. FEMS Microbiology Ecology, 96(6): fiaa083. https://doi.org/10.1093/femsec/fiaa083 |
| Reed, H. E., Martiny, J. B. H., 2013. Microbial Composition Affects the Functioning of Estuarine Sediments. The ISME Journal, 7(4): 868–879. https://doi.org/10.1038/ismej.2012.154 |
| Ren, L. J., Jeppesen, E., He, D., et al., 2015. pH Influences the Importance of Niche-Related and Neutral Processes in Lacustrine Bacterioplankton Assembly. Applied and Environmental Microbiology, 81(9): 3104–3114. https://doi.org/10.1128/aem.04042-14 |
| Roguet, A., Laigle, G. S., Therial, C., et al., 2015. Neutral Community Model Explains the Bacterial Community Assembly in Freshwater Lakes. FEMS Microbiology Ecology, 91(11): fiv125. https://doi.org/10.1093/femsec/fiv125 |
| Sadeghi, J., Chaganti, S. R., Shahraki, A. H., et al., 2021. Microbial Community and Abiotic Effects on Aquatic Bacterial Communities in North Temperate Lakes. Science of the Total Environment, 781: 146771. https://doi.org/10.1016/j.scitotenv.2021.146771 |
| Stegen, J. C., Lin, X. J., Konopka, A. E., et al., 2012. Stochastic and Deterministic Assembly Processes in Subsurface Microbial Communities. The ISME Journal, 6(9): 1653–1664. https://doi.org/10.1038/ismej.2012.22 |
| Stegen, J. C., Lin, X. J., Fredrickson, J. K., et al., 2015. Estimating and Mapping Ecological Processes Influencing Microbial Community Assembly. Frontiers in Microbiology, 6: 370. https://doi.org/10.3389/fmicb.2015.00370 |
| Stegen, J. C., Lin, X. J., Fredrickson, J. K., et al., 2013. Quantifying Community Assembly Processes and Identifying Features that Impose them. The ISME Journal, 7(11): 2069–2079. https://doi.org/10.1038/ismej.2013.93 |
| Tamaki, H., Wright, C. L., Li, X. Z., et al., 2011. Analysis of 16S rRNA Amplicon Sequencing Options on the Roche/454 Next-Generation Titanium Sequencing Platform. PLoS One, 6(9): e25263. https://doi.org/10.1371/journal.pone.0025263 |
| Tang, L. Y., Duan, X. F., Kong, F. J., et al., 2018. Influences of Climate Change on Area Variation of Qinghai Lake on Qinghai-Tibetan Plateau since 1980s. Scientific Reports, 8(1): 7331. https://doi.org/10.1038/s41598-018-25683-3 |
| Tang, X. M., Xie, G. J., Shao, K. Q., et al., 2021. Aquatic Bacterial Diversity, Community Composition and Assembly in the Semi-Arid Inner Mongolia Plateau: Combined Effects of Salinity and Nutrient Levels. Microorganisms, 9(2): 208. https://doi.org/10.3390/microorganisms9020208 |
| Tang, X. M., Xie, G. J., Shao, K. Q., et al., 2020. Contrast Diversity Patterns and Processes of Microbial Community Assembly in a River-Lake Continuum across a Catchment Scale in Northwestern China. Environmental Microbiome, 15(1): 10. https://doi.org/10.1186/s40793-020-00356-9 |
| Volkov, I., Banavar, J. R., Hubbell, S. P., et al., 2003. Neutral Theory and Relative Species Abundance in Ecology. Nature, 424(6952): 1035–1037. https://doi.org/10.1038/nature01883 |
| Waldrop, M. P., Balser, T. C., Firestone, M. K., 2000. Linking Microbial Community Composition to Function in a Tropical Soil. Soil Biology and Biochemistry, 32(13): 1837–1846. https://doi.org/10.1016/s0038-0717(00)00157-7 |
| Wang, B. C., Yang, J., Jiang, H. C., et al., 2019. Chemical Composition of n-Alkanes and Microbially Mediated n-Alkane Degradation Potential Differ in the Sediments of Qinghai-Tibetan Lakes with Different Salinity. Chemical Geology, 524: 37–48. https://doi.org/10.1016/j.chemgeo.2019.05.038 |
| Wang, J. J., Shen, J., Wu, Y. C., et al., 2013. Phylogenetic Beta Diversity in Bacterial Assemblages across Ecosystems: Deterministic Versus Stochastic Processes. The ISME Journal, 7(7): 1310–1321. https://doi.org/10.1038/ismej.2013.30 |
| Wang, W. D., Liu, W. Y., Wu, D., et al., 2019. Differentiation of Nitrogen and Microbial Community in the Littoral and Limnetic Sediments of a Large Shallow Eutrophic Lake (Chaohu Lake, China). Journal of Soils and Sediments, 19(2): 1005–1016. https://doi.org/10.1007/s11368-018-2090-4 |
| Williams, W. D., 1996. What Future for Saline Lakes?. Environment: Science and Policy for Sustainable Development, 38(9): 12–39. https://doi.org/10.1080/00139157.1996.9930999 |
| Willis, R. B., Montgomery, M. E., Allen, P. R., 1996. Improved Method for Manual, Colorimetric Determination of Total Kjeldahl Nitrogen Using Salicylate. Journal of Agricultural and Food Chemistry, 44(7): 1804–1807. https://doi.org/10.1021/jf950522b |
| Wu, Q. L., Zwart, G., Schauer, M., et al., 2006. Bacterioplankton Community Composition along a Salinity Gradient of Sixteen High-Mountain Lakes Located on the Tibetan Plateau, China. Applied and Environmental Microbiology, 72(8): 5478–5485. https://doi.org/10.1128/AEM.00767-06 |
| Xiong, J. B., Liu, Y. Q., Lin, X. G., et al., 2012. Geographic Distance and pH Drive Bacterial Distribution in Alkaline Lake Sediments across Tibetan Plateau. Environmental Microbiology, 14(9): 2457–2466. https://doi.org/10.1111/j.1462-2920.2012.02799.x |
| Xue, Y. Y., Chen, H. H., Yang, J. R., et al., 2018. Distinct Patterns and Processes of Abundant and Rare Eukaryotic Plankton Communities Following a Reservoir Cyanobacterial Bloom. The ISME Journal, 12(9): 2263–2277. https://doi.org/10.1038/s41396-018-0159-0 |
| Yang, J., Ma, L. A., Jiang, H. C., et al., 2016a. Salinity Shapes Microbial Diversity and Community Structure in Surface Sediments of the Qinghai-Tibetan Lakes. Scientific Reports, 6: 25078. https://doi.org/10.1038/srep25078 |
| Yang, J., Jiang, H. C., Wu, G., et al., 2016b. Distinct Factors Shape Aquatic and Sedimentary Microbial Community Structures in the Lakes of Western China. Frontiers in Microbiology, 7: 1782. https://doi.org/10.3389/fmicb.2016.01782 |
| Yang, J., Jiang, H. C., Liu, W., et al., 2018. Benthic Algal Community Structures and Their Response to Geographic Distance and Environmental Variables in the Qinghai-Tibetan Lakes with Different Salinity. Frontiers in Microbiology, 9: 578. https://doi.org/10.3389/fmicb.2018.00578 |
| Yang, J., Jiang, H. C., Dong, H. L., et al., 2019. A Comprehensive Census of Lake Microbial Diversity on a Global Scale. Science China Life Sciences, 62(10): 1320–1331. https://doi.org/10.1007/s11427-018-9525-9 |
| Yang, J., Jiang, H. C., Sun, X. X., et al., 2020. Minerals Play Key Roles in Driving Prokaryotic and Fungal Communities in the Surface Sediments of the Qinghai-Tibetan Lakes. FEMS Microbiology Ecology, 96(4): fiaa035. https://doi.org/10.1093/femsec/fiaa035 |
| Yang, J., Jiang, H. C., Sun, X. X., et al., 2021. Distinct Co-Occurrence Patterns of Prokaryotic Community between the Waters and Sediments in Lakes with Different Salinity. FEMS Microbiology Ecology, 97(1): fiaa234. https://doi.org/10.1093/femsec/fiaa234 |
| Yue, L. Y., Kong, W. D., Li, C. G., et al., 2021. Dissolved Inorganic Carbon Determines the Abundance of Microbial Primary Producers and Primary Production in Tibetan Plateau Lakes. FEMS Microbiology Ecology, 97(2): fiaa242. https://doi.org/10.1093/femsec/fiaa242 |
| Zhang, G. Q., Yao, T. D., Xie, H. J., et al., 2020. Response of Tibetan Plateau Lakes to Climate Change: Trends, Patterns, and Mechanisms. Earth-Science Reviews, 208: 103269. https://doi.org/10.1016/j.earscirev.2020.103269 |
| Zhang, G. Z., Wei, G. F., Wei, F. G., et al., 2021. Dispersal Limitation Plays Stronger Role in the Community Assembly of Fungi Relative to Bacteria in Rhizosphere across the Arable Area of Medicinal Plant. Frontiers in Microbiology, 12: 713523. https://doi.org/10.3389/fmicb.2021.713523 |
| Zhang, W. Z., Liu, Y. Q., Geng, M. D., et al., 2022. Extracellular Enzyme Stoichiometry Reveals Carbon and Nitrogen Limitations Closely Linked to Bacterial Communities in China's Largest Saline Lake. Frontiers in Microbiology, 13: 1002542. https://doi.org/10.3389/fmicb.2022.1002542 |
| Zhao, D. Y., Cao, X. Y., Huang, R., et al., 2017. The Heterogeneity of Composition and Assembly Processes of the Microbial Community between Different Nutrient Loading Lake Zones in Taihu Lake. Applied Microbiology and Biotechnology, 101(14): 5913–5923. https://doi.org/10.1007/s00253-017-8327-0 |
| Zheng, M. P., 1997. An Introduction to Saline Lakes on the Qinghai-Tibet Plateau. Springer, Dordrecht. 76. https://doi.org/10.1007/978-94-011-5458-1 |
| Zhou, J. Z., Ning, D. L., 2017. Stochastic Community Assembly: Does it Matter in Microbial Ecology?. Microbiology and Molecular Biology Reviews: MMBR, 81(4): e00002-17. https://doi.org/10.1128/MMBR.00002-17 |
| Zhou, J. Z., Liu, W. Z., Deng, Y., et al., 2013. Stochastic Assembly Leads to Alternative Communities with Distinct Functions in a Bioreactor Microbial Community. mBio, 4(2): e00584-12. https://doi.org/10.1128/mbio.00584-12 |
| Zhou, J. Z., Deng, Y., Zhang, P., et al., 2014. Stochasticity, Succession, and Environmental Perturbations in a Fluidic Ecosystem. Proceedings of the National Academy of Sciences of the United States of America, 111(9): E836–E845. https://doi.org/10.1073/pnas.1324044111 |
| Zhou, X. Q., Guo, Z. Y., Chen, C. R., et al., 2017. Soil Microbial Community Structure and Diversity are Largely Influenced by Soil pH and Nutrient Quality in 78-Year-Old Tree Plantations. Biogeosciences, 14(8): 2101–2111. https://doi.org/10.5194/bg-14-2101-2017 |
Figures(5) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: