| Citation: | Yang PU, Jun-hua HUANG, Xian-yu HUANG, Jing-wei CUI, Chao-yong HU. Acyclic Alkanes in the Soil over Heshang Cave in Qingjiang, Hubei Province. Journal of Earth Science, 2006, 17(2): 115-120.

|
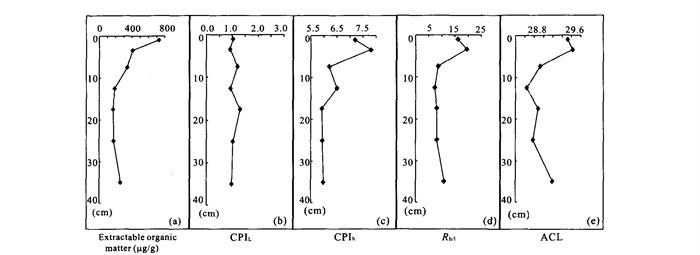
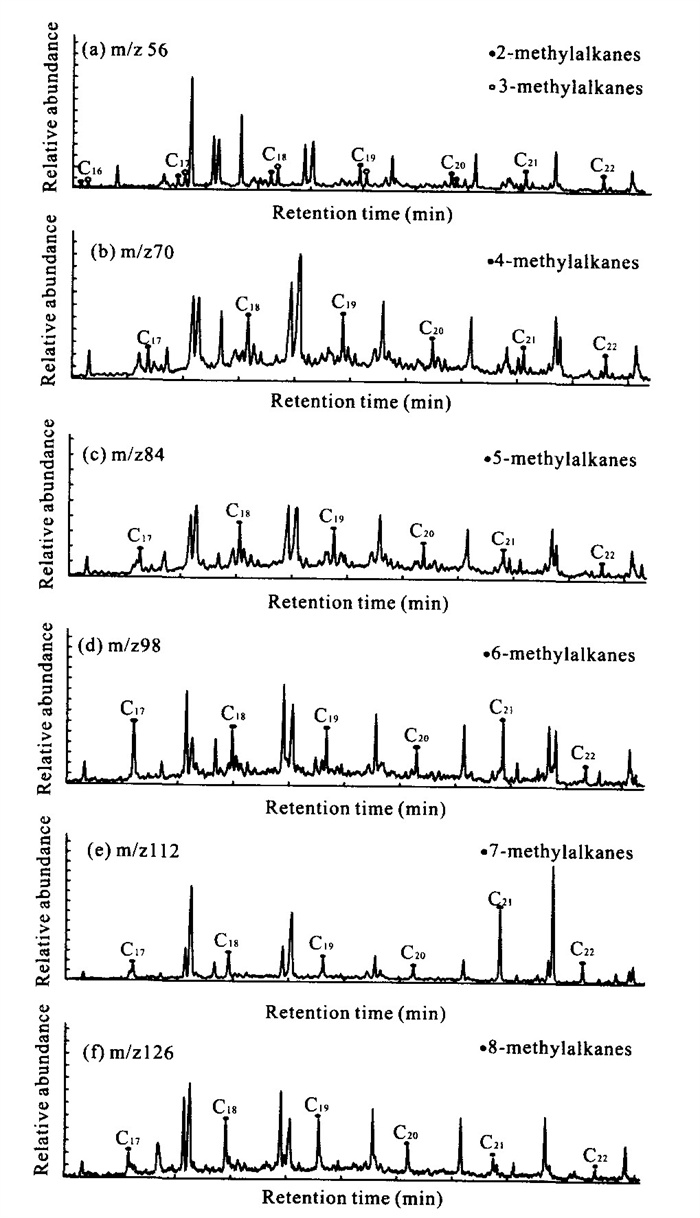
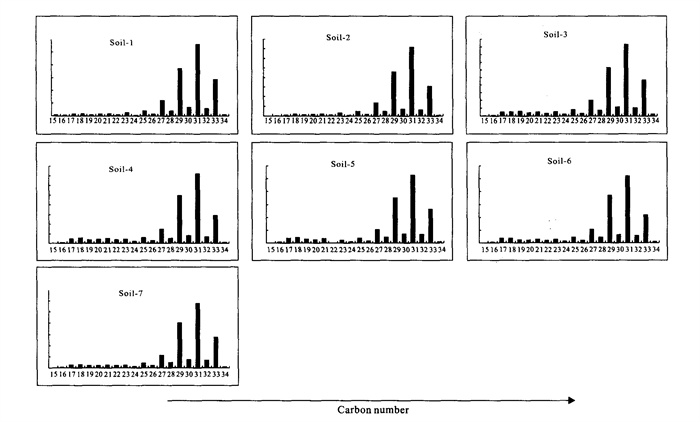
The reports that relate to the biomarker's fate and characteristics of the modern soil in the karst area are very lacking. By using gas chromatography-mass spectrometry (GC-MS), a series of biomarkers were identified from the soils collected over Heshang cave (和尚洞) in Qingjiang (清江), Hubei (湖北) Province. The distribution of n-alkanes is mainly from C_ 25 to C_ 33 in carbon number, with a maximum at C_ 31. They have a strong odd-over-even carbon number predominance. These characteristics represent an input mainly from higher plants. The lipid parameters, including CPIh (carbon preference index), R_ h/l (ratio of lower- to higher-molecular-weight homologues) and ACL (average chain length), show comparable trends with depth, probably reflecting vegetation change and microbial degradation. Series of monomethylalkanes and diploptene are present in the extractable organic matter; they might be derived from soil microbes, cyanobacteria in particular.

| Albro, P. W., 1976. Bacterial Waxes. In: Kolattukudy, PE., ed., Chemistry and Biochemistry of Natural WaxesElsevier, Amsterdam. 419 -445. |
| Baas, M., Pancost, R., Van Geel, B., et al., 2000. A Com-parative Study of Lipids in Sphagnum Species. Organic Geochemistry, 31: 535 -541. doi: 10.1016/S0146-6380(00)00037-1 |
| Bull, I. D., Van Bergen, P. F., Nott, C. J., et al., 2000 Organic Geochemical Studies of Soils fromthe Rothamsted Classical Experi ments—Ⅴ. The Fate of Lipids in Different Long-Term Experi ments. Organic Geochemistry 31: 389 -408. |
| Bull, I. D., Van Bergen, P. F., Poulton, P. R., et al., 1998. Organic Geochemical Studies of Soils from theRothamsted Classical Experi ments—Ⅱ. Soils from theHoosfield Spring Barley Experi ment Treated with Differ-ent Quantities of Manure. Organic Geochemistry, 28: 11-26. |
| Cranwell, P. A., Eglinton, G., Robinson, N., 1987. Lipidsof Aquatic Organisms as Potential Contributors to Lacus-trine Sedi ments. Organic Geochemistry. 11: 513 -527. doi: 10.1016/0146-6380(87)90007-6 |
| Dachs, J., Bayona, J. M., Fowler, S. W., et al., 1998. Ev-idence for Cyanobacterial Inputs and Heterotrophic Alter-ation of Lipids in Sinking Particles in the Alboran Sea (SW Mediterranean). Marine Chemistry, 60: 189 -201. doi: 10.1016/S0304-4203(97)00105-9 |
| De Rosa, M., Gambacorta, A., Minale, L., 1971. Bacterial Triterpenes. Chem. Comm., 619 -620. |
| Dembitsky, V. M., Dor, I., Shkrob, I., et al., 2001. Branched Alkanes and Other Apolar Compounds Pro-duced by Thecyanobacterium Microcoleus Vaginatus fromthe Negevdesert. Russian Journal of Bioorganic Chemis-try, 27: 110 -119. doi: 10.1023/A:1011385220331 |
| Gelpi, E., Schneider, H., Mann, J., et al., 1970. Hydro-carbons of Geochemical Significance in Microscopic Al-gae. Phytochemistry, 9: 603 -612. doi: 10.1016/S0031-9422(00)85700-3 |
| Gernot, A., Volker, T., Andreas, R., et al., 1999. Biofil mExopolymers Control Microbialite Formation at ThermalSprings Discharging into the Alkaline Pyramid Lake, Ne-vada, USA. Sedi mentary Geology, 126: 159 -176. doi: 10.1016/S0037-0738(99)00038-X |
| Gohring, K. E. H., Schenck, P. A., Engelhardt, E. D., 1967. A New Series of Isoprenoid Isoalkanes in CrudeOils and Cretaceous Bituminous Shale. Nature, 215: 503-505. doi: 10.1038/215503a0 |
| Han, J., Mccarthy, E. D., Calvin, M., et al., 1968. Hy-drocarbon Constituents of the Blue-Green Algae NostocMuscorum, Anacystis Nidulans, Phormidium Luridumand Chlorogloea Frischtii. Journal ofthe Chemical Soci-ety C, 2785 -2791. |
| Hendrix, P. F., Peterson, A. C., Beare, M. H., et al., 1998. Long-Term Effects of Earthworms on MicrobialBiomass Nitrogen in Coarse and Fine Textured Soils. Applied Soil Ecology, 9: 375 -380. doi: 10.1016/S0929-1393(98)00092-4 |
| Jackson, M. J., Powell, T. G., Summons, R. E., et al., 1986. Hydrocarbon Showand PetroleumSource Rocks inSedi ments as Old as 1.7×109Years. Nature, 322: 727-729. doi: 10.1038/322727a0 |
| Kenig, F., Dirk-Jan, H. S., David, C., 2005. Structure andDistribution of Branched Aliphatic Alkanes with Quater-nary Carbon Atoms in Cenomanian and Turonian BlackShales of Pasquia Hills (Saskatchewan, Canada). Organic Geochemistry, 36: 117 -138. doi: 10.1016/j.orggeochem.2004.06.014 |
| Kenig, F., Sinninghe, J. S., Dalen, A. C. K., et al., 1995. Occurrence and Origin of Monomethyl-Alkanes, Di meth-ylalkanes, and Tri methylalkanes in Modern and Holocene Cyanobacterial Mats from Abu-Dhabi, United-Arab-Emirates. Geochi micaet Cosmochi mica Acta, 59: 2999 -3015. doi: 10.1016/0016-7037(95)00190-5 |
| Koster, J., Volkman, J. K., Rullkotter, J., et al., 1999. Mono-, Di- and Tri methyl-Branched Alkanes in Culturesof the Filamentous Cyanobacterium Calothrix Scopulo-rum. Organic Geochemistry, 30: 1367 -1379. doi: 10.1016/S0146-6380(99)00110-2 |
| Liang, B., Xie, S. C., Gu, Y. S., et al., 2005. Distributionofn-Alkanes as Indicative of Paleovegetation Change inPleistocene Red Earth in Xuancheng, Anhui. EarthScience—Journal of China University of Geosciences, 30 (2): 129 -132 (in Chinese with English Abstract). |
| Lu, H., Peng, P. A., Sun, Y. G., 2003. Molecular andStable Carbon Isotopic Composition of Monomethylal-kanes from One Oil Sand Sample: Source I mplications. Organic Geochemistry, 34: 745 -754. doi: 10.1016/S0146-6380(03)00039-1 |
| Poynter, J. G., Farri mond, P., Robinson, N., et al., 1999. Aeolian-Derived Higher Plants Lipids in the Marine Sedi-mentary Record: Links with Paleocli mate. In: Leinen, M., Sarnthein, M., eds., Paleocli matology and Paleom-eteorology: Modern and Past Patterns of Global At mos-pheric Transport. Kluwer, Dordrecht. 435 -462. |
| Robinson, N., Eglinton, G., 1990. Lipid Chemistry of Ice-landichot Spring Microbial Mats. Organic Geochemistry, 15: 291 -298. doi: 10.1016/0146-6380(90)90007-M |
| Sheng, G. Y., Cai, K. Q., Yang, X. X., et al., 1999. Long-Chain Alkenones in Hotong Qagan Nurlake Sedi-ments and Its Paleocli matic I mplications. Chinese ScienceBulletin, 44 (3): 259 -263. |
| Shiea, J., Brassell, S. C., Ward, D. M., 1990. Mid-chainBranched Mono-and Di methyl Alkanes in Hot Spring Cy-anobacterial Mats: A Direct Biogenic Source for BranchedAlkanes in Ancient Sedi ments?Organic Geochemistry, 15: 223 -231. doi: 10.1016/0146-6380(90)90001-G |
| Shiea, J., Brassell, S. C., Ward, D. M., 1991. ComparativeAnalysis of Extractable Lipids in Hot Spring MicrobialMats and Their Component Photosynthetic Bacteria. Organic Geochemistry, 17: 309 -319. doi: 10.1016/0146-6380(91)90094-Z |
| Summons, R. E., Powell, T. G., Boreham, C. J., 1988. Petroleum Geology and Geochemistry of the Middle Prot-erozoic Mcarthur Basin, Northern Australia: Ⅲ Compo-sition of Extractable Hydrocarbons. Geochi micaet Cosmochi mica Acta, 52: 1747 -1763. doi: 10.1016/0016-7037(88)90001-4 |
| Thiel, V., Jenisch, A., Worheide, G., et al., 1999. Mid-chain Branched Alkanoic Acids from"Living Fossil"Demosponges: A Link to Ancient Sedi mentary Lipids?Organic Geochemistry, 30: 1 -14. doi: 10.1016/S0146-6380(98)00200-9 |
| Vinther, F. P., Eiland, F., Lind, L., 1999. Elsgaard Micro-bial Biomass and Numbers of Denitrifiers Related toMacropore Channels in Agricultural and Forest Soils. Soil Biology and Biochemistry, 31: 603 -611. doi: 10.1016/S0038-0717(98)00165-5 |
| Volkman, J. K., Barrett, S. M., Blackburn, S. I., et al., 1995. Alkenones in Gephyrocapsa Oceanica: I mplicationsfor Studies of Paleocli mate. Geochi mica et Cosmochi micaActa, 59: 513 -520. doi: 10.1016/0016-7037(95)00325-T |
| Wang, Z. Y., Liu, Z. H., Yi, Y., et al., 2003. Features ofLipids and Their Significance in Modern Soils from Vari-ous Cli mato-vegetation Regions. Acta Pedologica Sinica, 40 (6): 967 -970 (in Chinese with English Abstract). |
| Wright, C. J., Coleman, D. C., 2000. Cross-Site Comparisonof Soil Microbial Biomass, Soil Nutrient Status, andNematode Trophic Groups. Pedobiologia, 44: 2 -23. doi: 10.1078/S0031-4056(04)70024-4 |
| Xie, S. C., Huang, J. H., Wang, H. M., et al., 2005. TheSignificance of the Fatty Acids in the Stalagmite inHeshang Cave in Qingjiang, Hubei Province. Science in China (Series D), 35 (3): 246-251 (in Chinese). |
| Xie, S. C., Nott, C. J., Avsejs, L. A., et al., 2004. Mo-lecular and Isotopic Stratigraphyin an Ombrotrophic Mirefor Paleocli mate Reconstruction. Geochi mica et Cosmo-chi mica Acta, 68: 2849 -2862. doi: 10.1016/j.gca.2003.08.025 |
| Xie, S. C., Wang, Z. Y., Wang, H. M., et al., 2002. Grassy Vegetation since the Last Interglacial in the LoessPlateau: Molecular Fossil Record. Science in China (Se-ries D), 32 (1): 28 -35 (in Chinese). |
| Xie, S. C., Yao, T. D., Kang, S. C., et al., 1999. Cli maticand Environmental I mplications from Organic Matter inDasuopu Glacier in Xixiabangma in Qinghai-Tibetan Plat-eau. Science in China (Series D), 42 (4): 383 -391. |
| Xie, S. C., Yi, Y., Huang, J. H., et al., 2003a. Lipid Dis-tribution in a Subtropical Southern China Stalagmite: ARecord of Soil Ecosystem Response to Paleocli mateChange. Quaternary Research, 60: 340 -347 (in Chinesewith English Abstract). doi: 10.1016/j.yqres.2003.07.010 |
| Xie, S. C., Yi, Y., Liu, Y. Y., et al., 2003b. VermicularRed Earth in Response to the Global Cli matic Change inSouth China: Molecular Fossil Record. Science in China (Series D), 33 (5): 411 -417 (in Chinese). |
Figures(3) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: