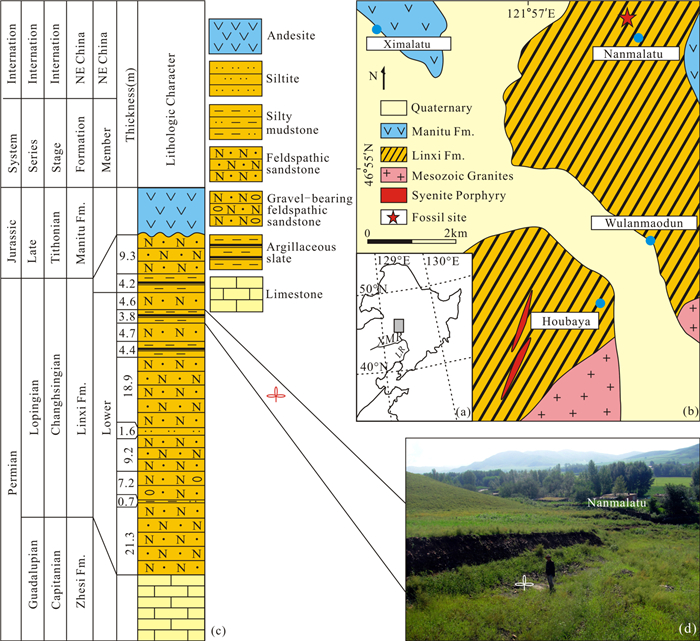
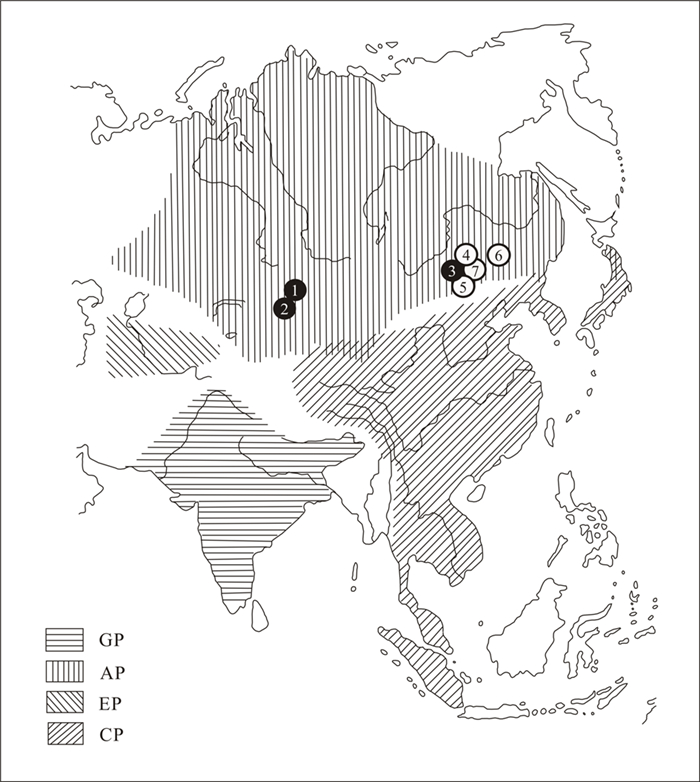
| Citation: | Zhen Tang, Yi Zhang, Serge V. Naugolnykh, Changqing Zheng, Lu Shi, Tao Qin, Junhao Huang. Ufadendron elongatum sp. nov., an Angaran Lycopsid from the Upper Permian of Inner Mongolia, China. Journal of Earth Science, 2020, 31(1): 1-8. doi: 10.1007/s12583-019-1230-0

|

| Archangelsky, S., Azcuy, C. L., Wagner, R. H., 1981. Three Dwarf Lycophytes from the Carboniferous of Argentina. Scripta Geology, 64: 1-35 |
| Baud, A., 2018. Final Results and Recommendations of the Last 10 Years IGCP 572 and 630 Field Workshops in South Turkey, Oman, India (Kashmir) and Armenia. Journal of Earth Science, 29(4): 733-744. https://doi.org/10.1007/s12583-018-0796-2 |
| Boucot, A. J, Chen, X., Scotese, C. R., et al., 2013. Phanerozoic Plaeoclimate: An Atlas of Lithologic Indicators of Climate. Society for Sedimentary Geology, Oklahoma. 478 |
| Chaloner, W. G., Leistikow, K. U., Hill, A., 1979. Brasilodendron gen. nov. and B. pedroanum (Carruthers) comb. nov., a Permian Lycopod from Brazil. Review of Palaeobotany and Palynology, 28(2): 117-136. https://doi.org/10.1016/0034-6667(79)90004-6 |
| Halle, T. G., 1937. The Relation between the Late Paleozoic Floras of Eastern and Northern Asia. C. R. Deuxième Congrès Pour L'Avancement des Etudes de Stratigraphie Du Carbonifère, 1: 237-245 |
| Huang, B. H., 1977. Permian Flora from the Southeastern Part of the Xiao Hinggan Ling (Lesser Khingan Mt.), NE China. Geological Publishing House, Bejing. 79 (in Chinese) |
| Huang, B. H., 1993. Carboniferous and Permian Systems and Floras in the Da Hinggan Range. Geological Publishing House, Bejing. 141 (in Chinese) |
| Khan, M. A., Bera, S., 2016. Occurrence of Persea Mill. from the Siwalik Forest of Darjeeling, Eastern Himalaya: Paleoclimatic and Paleogeographic Implications. Journal of Earth Science, 27(5): 883-890. https://doi.org/10.1007/s12583-016-0902-2 |
| Leven, E. J., Naugolnykh, S. V., Gorgij, M. N., 2011. New Findings of Permian Marine and Terrestrial Fossils in Central Iran (the Kalmard Block) and Their Significance for Correlation of the Tethyan, Uralian and West European Scales. RivistaItaliana di Paleontologia e Stratigrafia, 117(3): 355-374. https://doi.org/10.13130/2039-4942/5981 |
| Li, X. X., Shen, G. L., Tian, B. L., et al., 1995. Some Notes on Carboniferous and Permian Floras in China. In: Li, X. X., ed. Fossil Floras of China through the Geological Ages. Guangdong Science and Technology Press, Guangzhou. 244-302 |
| Meyen, S. V., 1972. Are There Ligula and Parichnos in Angara Carboniferous Lepidophytes?. Review of Palaeobotany and Palynology, 14(1/2): 149-157. https://doi.org/10.1016/0034-6667(72)90016-4 |
| Meyen, S. V., 1976. Carboniferous and Permian Lepidophytes of Angaraland. Palaeontographica Abt. B., 157: 112-157 |
| Naugolnykh, S. V., 2014. Fossil Flora from the Aleksandrovskoe Locality (Lower Permian, Kungurian; Krasnoufimsk District of the Sverdlovsk Region): Taxonomical Composition, Taphonomy, and a New Lycopsid Representative. Paleontological Journal, 48(2): 209-217. https://doi.org/10.1134/s0031030114020105 |
| Naugolnykh, S. V., 2016. Flora Permica. Plant World of Permian Period. Geos Press, Moscow. 336 (in Russian) |
| Naugolnykh, S. V., Uranbileg, L., 2018. A New Discovery of Glossopteris in Southeastern Mongolia as an Argument for Distant Migration of Gondwanan Plants. Journal of Asian Earth Sciences, 154: 142-148. https://doi.org/10.1016/j.jseaes.2017.11.039 |
| Radczenko, G. P., 1955. Index-Fossils of the Upper Palaeozoic Flora of the Sayan-Altai Region. Atlas Rukovod. Form Iskop. Fauny i Flory Zap. Sibiri, 2: 42-153 |
| Sun, K. Q., Cui, J. Z., Wang, S. J., 2010. Fossil flora of China, Volume 2: Fossil Pteridophytes in China. Higher Education Press, Bejing. 438 (in Chinese) |
| Sun, Y. W., Ding, H. S., Liu, H., et al., 2016. Fossil Plants from the Guadalupian Yujiabeigou Formation in the North Margin of North China Plate and Their Tectonic Implications. Journal of Jilin University (Earth Science Edition), 46(5): 1268-1283 (in Chinese with English Abstract) http://d.old.wanfangdata.com.cn/Periodical/cckjdxxb201605001 |
| Taylor, T. N., Taylor, E. L., Krings, M., 2009. Paleobotany-The Biology and Evolution of Fossil Plants. Academic Press (Elsevier), Boston. 1230 |
| Thomas, B. A., Meyen, S. V., 1984. A System of Form-Genera for the Upper Palaeozoic Lepidophyte Stems Represented by Compression-Impression Material. Review of Palaeobotany and Palynology, 41(3/4): 273-281. https://doi.org/10.1016/0034-6667(84)90049-6 |
| Wang, X. F., Chen, X. H., 2015. Stratigraphic Division and Correlation of Each Geological Period in China. Geological Publishing House, Beijing. 596 (in Chinese) |
| Zalessky, M. D., 1918. Flore Paléozoique de la Série d'Angara. Atlas. Mémoires du Comité Géologique: Nouvelle Série, 147: 5-76 |
| Zalessky, M. D., 1936. Sur Quelques Plantes Nouvelles du Système Anthracolitique du Bassin de Kousnetzk. Problems of Paleontology, Moscow University, 1: 223-236 |
| Zhang, Y. S., Niu, S. W., Tian, S. G., et al., 2012. The Discovery of Conchostracan Fossils in the Upper Permian Linxi Formation of Linxi Area, Inner Mongolia, and Its Geological Significance. Geological Bulletin of China, 31(9): 1394-1403 (in Chinese with English Abstract) http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=zgqydz201209004 |
| Zhang, Y., Wang, J., Wu, X. Y., 2006. Cathaysiodendron Yangshanense sp. nov. from the Early Carboniferous Yangshan Formation of Gushi, Henan. Acta Palaeontology Sinica, 45(2): 265-267 (in Chinese with English Abstract) http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=gswxb200602011 |
| Zhang, Y., Zheng, S. L., Naugolnykh, S. V., 2012. A New Species of Lepidopteris Discovered from the Upper Permian of China with Its Stratigraphic and Biologic Implications. Chinese Science Bulletin, 57(27): 3603-3609. https://doi.org/10.1007/s11434-012-5282-0 |

Figures(6) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: