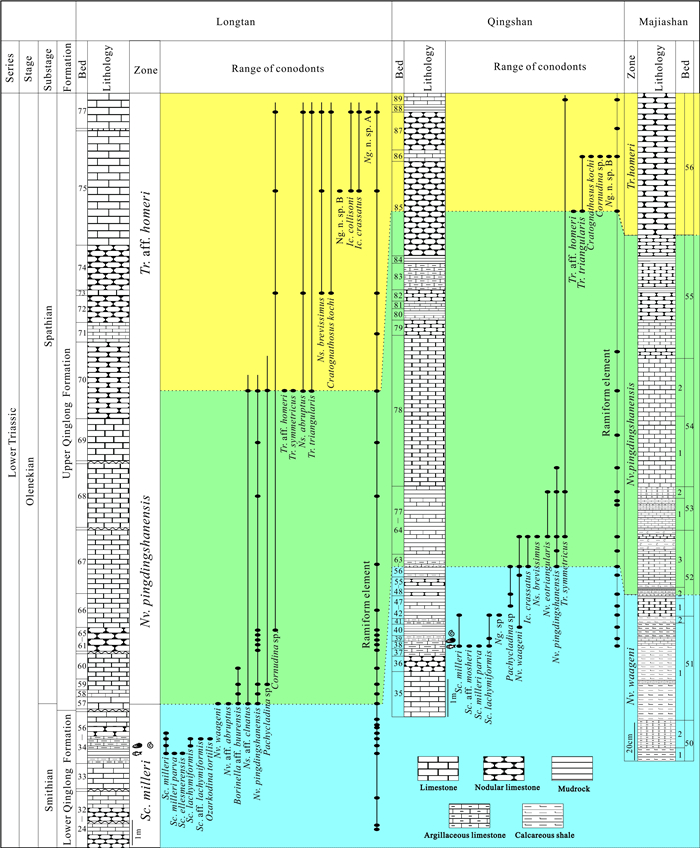
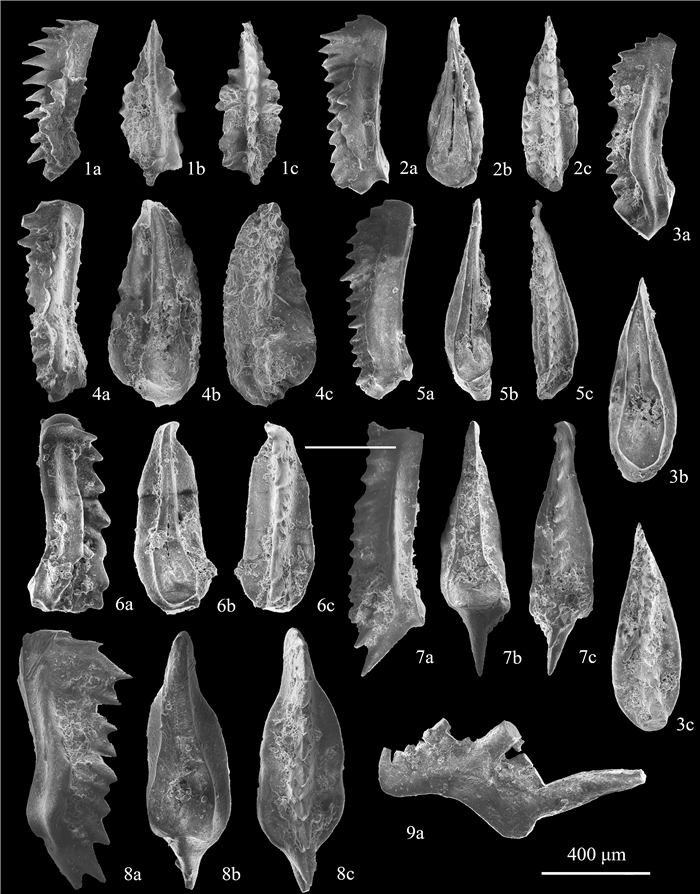
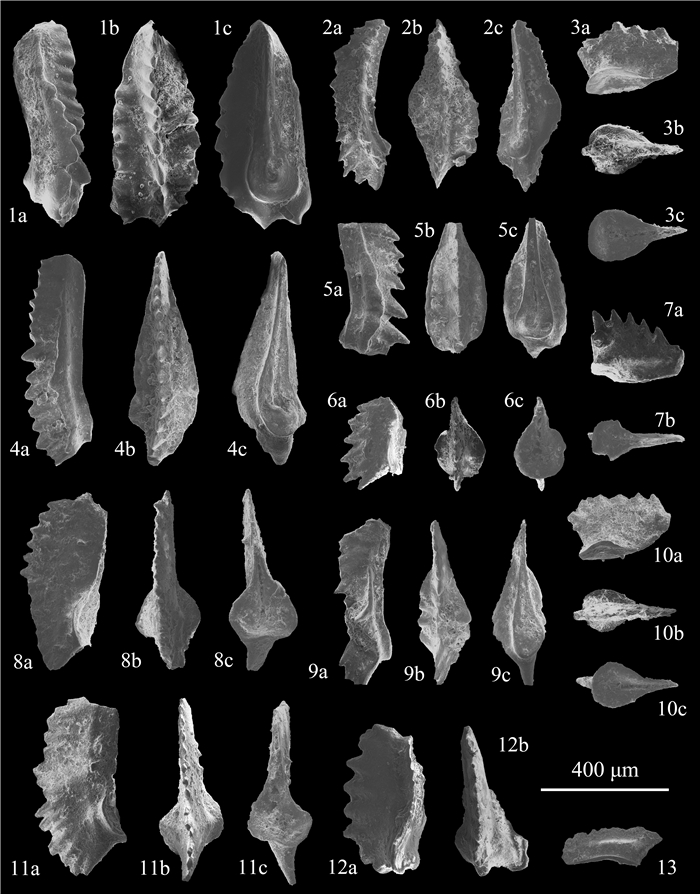
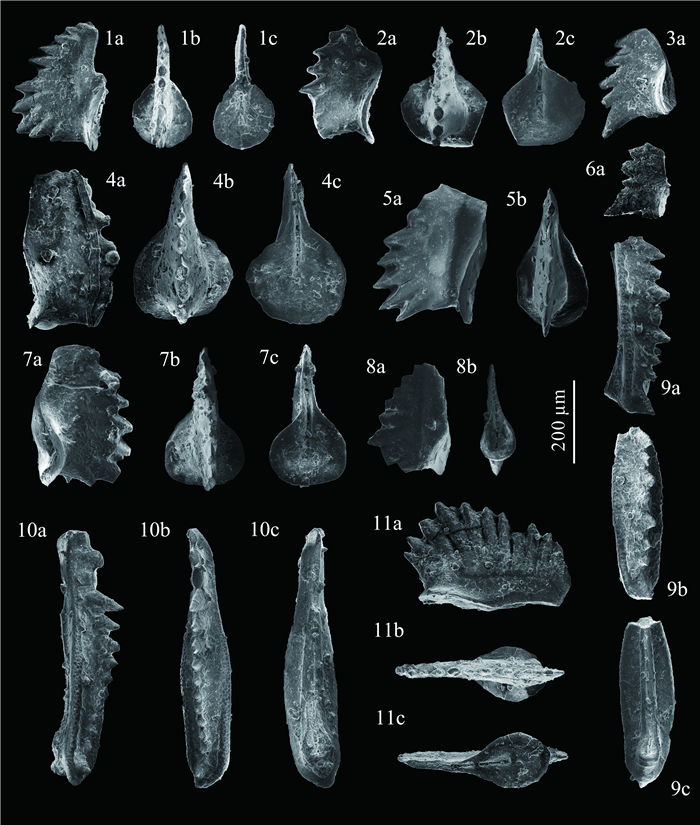
| Citation: | Shuang Liu, Zuoyu Sun, Cheng Ji, Min Zhou, Dayong Jiang. Conodont Biostratigraphy and Age of the Early Triassic Fish-Bearing-Nodule Levels from Nanjing and Jurong, Jiangsu Province, South China. Journal of Earth Science, 2020, 31(1): 9-22. doi: 10.1007/s12583-019-1232-y

|

| Bender, H., 1970. Zur Gliederung der Mediterranen Trias Ⅱ. Die Conodontenchronologie der Mediterranen Trias. Annales Géologiques des Pays Helléniques, 19: 465-540 (in French) |
| Chen, C. Z., Wang, Y. G., Wang, Z. H., et al., 1988. Triassic Biostratigraphy in Southern Jiangsu Province. In: Academy of Geological Sciences, Jiangsu Bureau of Petroleum, Prospecting and Nanjing Institute of Geology and Paleontology, Academia Sinica Sinian-Triassic Biostratigraphy of the Lower Yangtze Peneplatform in Jiangsu Region. Nanjing University Press, Nanjing (in Chinese) |
| Chen, M. J., 1996. Conodonts of Upper Permian-Lower Triassic of Zhenjiang Area. Acta Palaeontologica Sinica, 35: 430-441 (in Chinese with English Abstract) http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=QK199600128564 |
| Chen, Y. L., Jiang, H. S., Lai, X. L., et al., 2015. Early Triassic Conodonts of Jiarong, Nanpanjiang Basin, Southern Guizhou Province, South China. Journal of Asian Earth Sciences, 105: 104-121. https://doi.org/10.1016/j.jseaes.2015.03.014 |
| Chen, Y. L., Kolar-Jurkovšek, T., Jurkovšek, B., et al., 2016. Early Triassic Conodonts and Carbonate Carbon Isotope Record of the Idrija-Žiri Area, Slovenia. Palaeogeography, Palaeoclimatology, Palaeoecology, 444: 84-100. https://doi.org/10.13039/501100004329 |
| Chhabra, N. L., Kumar, S., 1992. Late Scythian through Early Carnian Conodont Assemblages and Their Biostratigraphic Significance from off Shore Carbonates of Northern Kumaun, Tethys Himalaya, India. Revue de Micropaleontologie, 35: 3-21 |
| Chhabra, N. L., Sahni, A., 1981. Late Lower Triassic and Early Triassic Conodont Faunas from Kashmir and Kumaun Sequences in Himalaya. Journal of the Palaeontological Society of India, 25: 47-135 |
| Clark, D. L., 1959. Conodonts from the Triassic of Nevada and Utah. Journal of Paleontology, 33: 305-312 |
| Fang, Q., Zhang, H. J., Wang, X. L., et al., 2015. The Discovery of Early Triassic Conodont in the Huashixia Area of Maduo County, Qinghai Province, NW China and Its Significance. Acta Micropalaeontologica Sinica, 30: 423-425, 441 (in Chinese with English Abstract) http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=wtgswxb201304009 |
| Feng, Z. Z., Bao, Z. D., Li, S. W., 1997. Lithofaceies Paleogeography of Early and Middle Triassic of South China. Petroleum Industry Press, Beijing (in Chinese) |
| Friedman, M., 2015. The Early Evolution of Ray-Finned Fishes. Palaeontology, 58(2): 213-228. https://doi.org/10.13039/501100004789 |
| Fuchs, G., Mostler, H., 1969. Mikrofaunen aus der Tibet-Zone, Himalaya. Verhandlungen der Geologischen Bundesanstalt, 133-145 |
| Gaetani, M., Jacobshagen, V., Nicora, A., et al., 1992. The Early-Middle Triassic Boundary at Chios (Greece). Rivista Italiana di Paleontologia e Stratigrafia, 98: 181-204 http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=HighWire000006031555 |
| Henderson, C., 1997. Uppermost Permian Conodonts and the Permian-Triassic Boundary in the Western Canada Sedimentary Basin. Bulletin of Canadian Petroleum Geology, 45: 693-707 |
| Ji, W. T., Tong, J. N., Zhao, L. S., et al., 2011. Lower-Middle Triassic Conodont Biostratigraphy of the Qingyan Section, Guizhou Province, Southwest China. Palaeogeography, Palaeoclimatology, Palaeoecology, 308(1/2): 213-223. https://doi.org/10.1016/j.palaeo.2010.08.020 |
| Jiang, W., Luo, Y. Q., Lu, T. Q., et al., 2000. The Lower Triassic Conodonts and Its Significance to Oil and Gas Exploration in Sichuan Basin. Acta Micropalaeontology Sinica, 17: 99-109 http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=wtgswxb200001010 |
| Jin, F., 2006. An Overview of Triassic Fishes from China. Vertebrata PalAsiatica, 44: 28-42 (in Chinese with English Abstract) http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=gjzdwxb200601002 |
| Jin, F., Wang, N. Z., Cai, Z. Q., 2003. A Revision of the Perleidid Fishes from the Lower Yangtze Region of South China. Vertebrata PalAsiatica, 7: 169-84 (in Chinese with English Abstract) http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=gjzdwxb200303001 |
| Kogan, I., Romano, C., 2016. Redescription of Saurichthys Madagascariensis Piveteau, 1945 (Actinopterygii, Early Triassic), with Implications for the Early Saurichthyid Morphotype. Journal of Vertebrate Paleontology, 36(4): e1151886. https://doi.org/10.1080/02724634.2016.1151886 |
| Komatsu, T., Takashima, R., Shigeta, Y., et al., 2016. Carbon Isotopic Excursions and Detailed Ammonoid and Conodont Biostratigraphies around Smithian-Spathian Boundary in the Bac Thuy Formation, Vietnam. Palaeogeography, Palaeoclimatology, Palaeoecology, 454: 65-74. https://doi.org/10.13039/501100001691 |
| Kozur, H., 2003. Integrated Ammonoid, Conodont and Radiolarian Zonation of the Triassic and Some Remarks to Stage/Substage Subdivision and the Numeric Age of the Triassic Stages. Albertiana, 28: 57-74 |
| Krystyn, L., 2005. A Revised Lower Triassic Intercalibrated Ammonoid-Conodont Time Scale of the Eastern Tethys Realm Based on Himalayan Data. Albertiana, 33: 53-54 |
| Krystyn, L., Bhargava, O. N., Richoz, S., 2007. A Candidate GSSP for the Base of the Olenekian Stage: Mud at Pin Valley; District Lahul and Spiti, Himachal Pradesh (Western Himalaya), India. Albertiana, 35: 4-29 |
| Leu, M., Bucher, H., Goudemand, N., 2018. Clade-Dependent Size Response of Conodonts to Environmental Changes during the Late Smithian Extinction. Earth-Science Reviews. https://doi.org/10.1016/j.earscirev.2018.11.003 |
| Li, Q. G., 2009. A New Parasemionotid-Like Fish from the Lower Triassic of Jurong, Jiangsu Province, South China. Palaeontology, 52(2): 369-384. https://doi.org/10.1111/j.1475-4983.2009.00848.x |
| Liang, D., Tong, J. N., Zhao, L. S., 2011. Lower Triassic Smithian-Spathian Boundary at West Pingdingshan Section in Chaohu, Anhui Province. Science China Earth Sciences, 54(3): 372-379. https://doi.org/10.1007/s11430-010-4145-2 |
| Liang, L., Tong, J. N., Song, H. J., et al., 2016. Lower-Middle Triassic Conodont Biostratigraphy of the Mingtang Section, Nanpanjiang Basin, South China. Palaeogeography, Palaeoclimatology, Palaeoecology, 459: 381-393. https://doi.org/10.1016/j.palaeo.2016.07.027 |
| Liu, G. B., Feng, H. Z., Wang, J. X., et al., 2002. Early Triassic Fishes from Jurong, Jiangsu. Acta Paleontologica Sinica, 41: 27-52 (in Chinese with English Abstract) http://d.old.wanfangdata.com.cn/Periodical/gswxb200201005 |
| Marramà, G., Lombardo, C., Tintori, A., Carnevale, G., 2017. Redescription of 'Perleidus' (Osteichthyes, Actinopterygii) from the Early Triassic of Northwestern Madagascar. Rivista Italiana di Paleontologia e Stratigrafia, 123(2): 219-242 http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=ddf28e1da5a5656a708f2705c9bcfd3c |
| Mosher, L. C., 1973. Triassic Conodonts from British Columbia and the Northern Arctic Islands. Bulletin of the Geological Survey of Canada, 222: 141-192 |
| Müller, K. J., 1956. Triassic Conodonts from Nevada. Journal of Paleontology, 30: 818-830 http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=3588c9912751fb6e90a61c452dcf55bf |
| Nakrem, H. A., Orchard, M. J., Weitschat, W., et al., 2008. Triassic Conodonts from Svalbard and Their Boreal Correlations. Polar Research, 27(3): 523-539. https://doi.org/10.3402/polar.v27i3.6190 |
| Nogami, Y., 1968. Trias-Conodonten von Timor, Malaysien und Japan. Memoirs of the Faculty of Science, Kyoto University, 34: 115-136 |
| Orchard, M. J., 1995. Taxonomy and Correlation of Lower Triassic (Spathian) Segminate Conodonts from Oman and Revision of Some Species of Neospathodus. Journal of Paleontology, 69(1): 110-122. https://doi.org/10.1017/s0022336000026962 |
| Orchard, M. J., 2008. Lower Triassic Conodonts from the Canadian Arctic, Their Intercalibration with Ammonoid-Based Stages and a Comparison with Other North American Olenekian Faunas. Polar Research, 27(3): 393-412. https://doi.org/10.3402/polar.v27i3.6187 |
| Orchard, M. J., Tozer, E. T., 1997. Triassic Conodont Biochronology, Its Calibration with the Ammonoid Standard, and a Biostratigraphic Summary for the Western Canada Sedimentary Basin. Bulletin of Canadian Petroleum Geology, 45(4): 675-692 |
| Orchard, M. J., Zonneveld, J. P., et al., 2009. The Lower Triassic Sulphur Mountain Formation in the Wapiti Lake Area: Lithostratigraphy, Conodont Biostratigraphy, and a New Biozonation for the Lower Olenekian (Smithian) Earth Science Sector (ESS) Contribution 20080714. Canadian Journal of Earth Sciences, 46(10): 757-790. https://doi.org/10.1139/e09-051 |
| Orchard, M. J., Lehrmann, D. J., Wei, J., et al., 2007a. Conodont from the Olenekian-Anisian Boundary Beds, Guandao, Guizhou Province, China. In: Lucas, S. G., Spielmann, J. A. eds., The Global Triassic. New Mexico Museum of Natural History and Science Bulletin, 41: 347-354 |
| Orchard, M. J., Gradinaru, E., Nicora, A., 2007b. A Summary of the Conodont Succession around the Olenekian-Anisian Boundary at Desli Caira, North Dobrogea, Romania. In: Lucas, S. G., Spielmann, J. A., eds., The Global Triassic. New Mexico Museum of Natural History and Science Bulletin, 41: 341-346 |
| Qian, M. P., Zhu, S. P., Zhao, F. M., et al., 1997. Discovery of Early Triassic Fish Fossils and Its Significances in Jurong, Jiangsu Province. Jiangsu Geology, 21: 65-71 (in Chinese with English Abstract) |
| Romano, C., Koot, M. B., Kogan, I., et al., 2014. Permian-Triassic Osteichthyes (Bony Fishes): Diversity Dynamics and Body Size Evolution. Biological Reviews, 91(1): 106-147. https://doi.org/10.13039/501100001711 |
| Scheyer, T. M., Romano, C., Jenks, J., et al., 2014. Early Triassic Marine Biotic Recovery: The Predators' Perspective. Plos ONE, 9(3): e88987. https://doi.org/10.1371/journal.pone.0088987 |
| Solien, M. A., 1979. Conodont Biostratigraphy of the Lower Triassic Thaynes Formation, Utah. Journal of Paleontology, 53: 276-306 |
| Su, D. Z., 1981. A New Species of Perleidus from Anhui. Vertebrata PalAsiatica, 19(2): 107-112 (in Chinese with English Abstract) |
| Sweet, W. C., 1970. Uppermost Permian and Lower Triassic Conodonts of the Salt Range and Trans-Indus Ranges, West Pakistan. In: Kummel, B., Teichert, C., eds., Stratigraphic Boundarty Problems: Permian and Triassic of West Pakistan. University of Kansas, Department of Geology, Special Publication, 4: 207-275 |
| Sweet, W. C., Mosher, L. C., Clark, D. L., 1971. Conodont Biostratigraphy of the Triassic. Geological Society of America Memoir, 127: 441-465 doi: 10.1016-S1367-9120(02)00021-4/ |
| Tian, C. R., 1982. Triassic Conodonts in the Tulong Section from Nyalam County, Xizang (Tibet), China. Geological Review, 3: 153-165 (in Chinese with English Abstract) |
| Tintori, A., Hitij, T., Jiang, D. Y., et al., 2014. Triassic Actinopterygian Fishes: The Recovery after the End-Permian Crisis. Integrative Zoology, 9(4): 394-411. https://doi.org/10.1111/1749-4877.12077 |
| Tong, J. N., Zakharov, Y. D., Wu, S. B., 2004. Early Triassic Ammonoid Succession in Chaohu, Anhui Provinve. Acta Palaeontologica Sinica, 43: 192-204 (in Chinese with English Abstract) |
| Tong, J. N., Zhou, X. G., Erwin, D. H., et al., 2006. Fossil Fishes from the Lower Triassic of Majiashan, Chaohu, Anhui Province, China. Journal of Paleontology, 80(1): 146-161. https://doi.org/10.1666/0022-3360(2006)080[0146:ffftlt]2.0.co; 2 doi: 10.1666/0022-3360(2006)080[0146:ffftlt]2.0.co;2 |
| Wang, H. M., Wang, X. L., Li, R. X., et al., 2005. Triassic Conodont Succession and Stage Subdivision of the Guandao Section, Bianyang, Luodian, Guizhou. Acta Palaeontologica Sinica, 44(4): 611-626 (in Chinese with English Abstract) http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=gswxb200504012 |
| Wang, H. Z., Shi, X. Y., Wang, X. L., et al., 2000. Research on the Sequence Stratigraphy of China. Guangdong Science and Technology Press, Guangzhou. 457 (in Chinese with English Abstract) |
| Wang, Z. H., Cao, Y. Y., 1981. Early Triassic Conodonts from Lichuan, Western Hubei. Acta Micropalaeontology Sinica, 20: 363-375 (in Chinese with English Abstract) http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=9763e2d3290c15cb61cd577eccfe60c3 |
| Wang, Z. H., Zhong, D., 1994. Triassic Conodonts from Different Facies in Eastern Yunnan, Western Guizhou and Northern Guangxi. Acta Micropalaeontologica Sinica, 11: 379-412 (in Chinese with English Abstract) |
| Wu, G. C., Yao, J. X., Ji, Z. S., 2007. Triassic Conodont Biostratigraphy in the Coqen Area, Western Gangdise, Tibet, China. Geological Bulletin of China, 26: 938-946 (in Chinese with English Abstract) |
| Yan, C. B., Wang, L. N., Jiang, H. S., 2013. Uppermost Permian to Lower Triassic Conodonts at Bianyang Section, Guizhou Province, South China. Palaios, 28(8): 509-522. https://doi.org/10.2110/palo.2012.p12-077r |
| Yang, S. R., Chu, Q. C., 1992. Study on Conodonts from Triassic Yongningzhen Formation, Southwestern Guizhou Province with a Discussion on Lower/Middle Triassic Boundary. Acta Scientiarum Naturalium Universitatis Pekinensis, 28: 723-732 (in Chinese with English Abstract) |
| Zhao, L. S., Chen, Y. L., Chen, Z. Q., et al., 2013. Uppermost Permian to Lower Triassic Conodont Zonation from Three Gorges Area, South China. Palaios, 28(8): 523-540. https://doi.org/10.2110/palo.2012.p12-107r |
| Zhao, L. S., Orchard, M. J., 2007. Lower Triassic Conodont Sequence in Chaohu, Anhui Province, China and Its Global Correlation. Paleogeography, Palaeoclimatology, Palaeoecology, 252: 24-38. https://doi.org/10.1016/j.palaeo.2006.11.032 |
| Zhao, L. S., Orchard, M. J., Tong, J. N., et al., 2007. Lower Triassic Conodont Sequence in Chaohu, Anhui Province, China and Its Global Correlation. Palaeogeography, Palaeoclimatology, Palaeoecology, 252(1/2): 24-38. https://doi.org/10.1016/j.palaeo.2006.11.032 |
| Zhao, L. S., Tong, J. N., Sun, Z. M., et al., 2008. A Detailed Lower Triassic Conodont Biostratigraphy and Its Implications for the GSSP Candidate of the Induan-Olenekian Boundary in Chaohu, Anhui Province. Progress in Natural Science, 18(1): 79-90. https://doi.org/10.1016/j.pnsc.2007.07.001 |
| Zhang, L., Orchard, M. J., Brayard, A., et al., 2019. The Smithian-Spathian Boundary (Late Early Triassic): A Review of Ammonoid, Conodont, and Carbon-Isotopic Criteria. Earth-Science Reviews. https://doi.org/10.13039/501100004701 |
| Zuo, J. X., Tong, J. N., Qiu, H. O., et al., 2006. Carbon Isotope Composition of the Lower Triassic Marine Carbonates, Lower Yangtze Region, South China. Science in China Series D: Earth Science, 49(3): 225-241. https://doi.org/10.1007/s11430-006-0225-8 |
Figures(7) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: