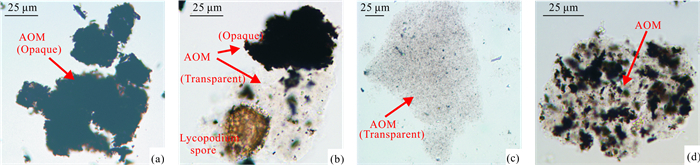
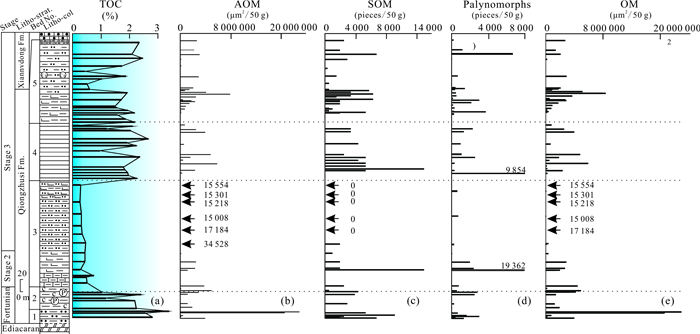
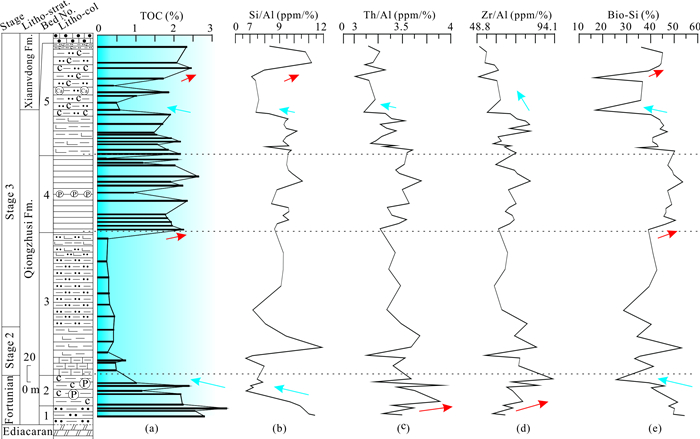
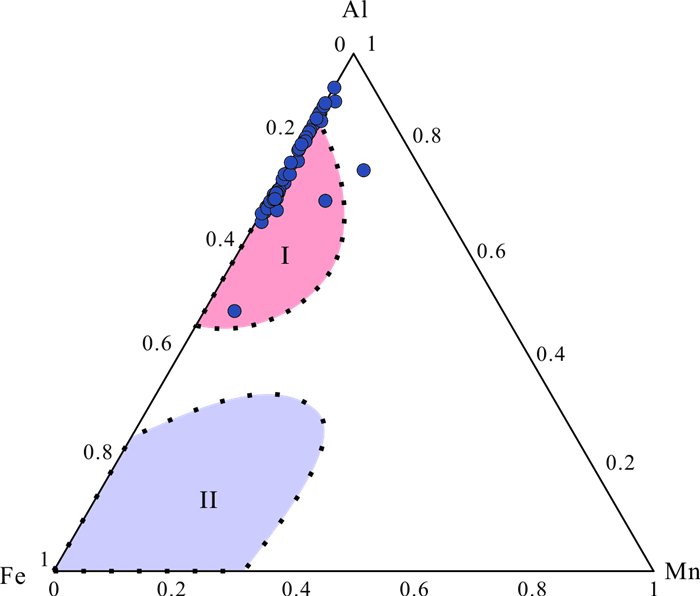
| Citation: | Shucan Zheng, Qinglai Feng, Nicolas Tribovillard, Thomas Servais, Yan Zhang, Bo Gao. New Insight into Factors Controlling Organic Matter Distribution in Lower Cambrian Source Rocks: A Study from the Qiongzhusi Formation in South China. Journal of Earth Science, 2020, 31(1): 181-194. doi: 10.1007/s12583-019-1240-y

|

| Adachi, M., Yamamoto, K., Sugisaki, R., 1986. Hydrothermal Chert and Associated Siliceous Rocks from the Northern Pacific Their Geological Significance as Indication Od Ocean Ridge Activity. Sedimentary Geology, 47(1/2): 125-148. https://doi.org/10.1016/0037-0738(86)90075-8 |
| Algeo, T. J., Lyons, T. W., 2006. Mo-Total Organic Carbon Covariation in Modern Anoxic Marine Environments: Implications for Analysis of Paleoredox and Paleohydrographic Conditions. Paleoceanography, 21(1): 1-23. https://doi.org/10.1029/2004pa001112 |
| Algeo, T. J., Maynard, J. B., 2004. Trace-Element Behavior and Redox Facies in Core Shales of Upper Pennsylvanian Kansas-Type Cyclothems. Chemical Geology, 206(3/4): 289-318. https://doi.org/10.1016/j.chemgeo.2003.12.009 |
| Algeo, T. J., Tribovillard, N., 2009. Environmental Analysis of Paleoceanographic Systems Based on Molybdenum-Uranium Covariation. Chemical Geology, 268(3/4): 211-225. https://doi.org/10.13039/100000001 |
| Arsairai, B., Wannakomol, A., Feng, Q. L., et al., 2016. Paleoproductivity and Paleoredox Condition of the Huai Hin Lat Formation in Northeastern Thailand. Journal of Earth Science, 27(3): 350-364. https://doi.org/10.1007/s12583-016-0666-8 |
| Banahan, S., Goering, J. J., 1986. The Production of Biogenic Silica and Its Accumulation on the Southeastern Bering Sea Shelf. Continental Shelf Research, 5(1/2): 199-213. https://doi.org/10.1016/0278-4343(86)90015-4 |
| Batten, D. J., 1996. Palynofacies and Palaeoenvironmental Interpretation. Journal of Micropalaeontology, 3: 1011-1064 |
| Batten, S. D., Freeland, H. J., 2007. Plankton Populations at the Bifurcation of the North Pacific Current. Fisheries Oceanography, 16(6): 536-546. https://doi.org/10.1111/j.1365-2419.2007.00448.x |
| Bhattacharya, S., Dutta, S., 2015. Neoproterozoic-Early Cambrian Biota and Ancient Niche: A Synthesis from Molecular Markers and Palynomorphs from Bikaner-Nagaur Basin, Western India. Precambrian Research, 266: 361-374. https://doi.org/10.1016/j.precamres.2015.05.029 |
| Braun, A., Chen, J., Waloszek, D., et al., 2007. First Early Cambrian Radiolaria. Geological Society, London, Special Publications, 286(1): 143-149. https://doi.org/10.1144/sp286.10 |
| Brooks, S. J., Harman, C., Hultman, M. T., et al., 2015. Integrated Biomarker Assessment of the Effects of Tailing Discharges from an Iron Ore Mine Using Blue Mussels (Mytilus Spp.). Science of the Total Environment, 524-525: 104-114. https://doi.org/10.1016/j.scitotenv.2015.03.135 |
| Burdige, D. J., 2006. Data Report: Dissolved Carbohydrates in Interstitial Waters from the Equatorial Pacific and Peru Margin. Proceedings of the Ocean Drilling Program. Scientific Results, 201: 1-10. https://doi.org/10.2973/odp.proc.sr.201.118.2006 |
| Burdige, D. J., 2007. Preservation of Organic Matter in Marine Sediments: Controls, Mechanisms, and an Imbalance in Sediment Organic Carbon Budgets?. Chemical Reviews, 107 (2): 467-485. https://doi.org/10.1021/cr050347q |
| Butterfield, N. J., 2007. Macroevolution and Macroecology through Deep Time. Palaeontology, 50(1): 41-55. https://doi.org/10.1111/j.1475-4983.2006.00613.x |
| Butterfield, N. J., 2011. Animals and the Invention of the Phanerozoic Earth System. Trends in Ecology & Evolution, 26(2): 81-87. https://doi.org/10.1016/j.tree.2010.11.012 |
| Cao, W., 2014. Cambrian Series 2 Shuijingtuo Formation Radiolarian Fauna from Zigui: [Dissertation]. China University of Geosciences, Wuhan. 49 (in Chinese with English Abstract) |
| Caron, D. A., Michaels, A. F., Swanberg, N. R., et al., 1995. Primary Productivity by Symbiont-Bearing Planktonic Sarcodines (Acantharia, Radiolaria, Foraminifera) in Surface Waters near Bermuda. Journal of Plankton Research, 17(1): 103-129. https://doi.org/10.1093/plankt/17.1.103 |
| Chang, S., Feng, Q. L., Clausen, S., et al., 2017. Sponge Spicules from the Lower Cambrian in the Yanjiahe Formation, South China: The Earliest Biomineralizing Sponge Record. Palaeogeography, Palaeoclimatology, Palaeoecology, 474: 36-44. https://doi.org/10.1016/j.palaeo.2016.06.032 |
| Chang, S., Clausen, S., Zhang, L., et al., 2018. New Probable Cnidarian Fossils from the Lower Cambrian of the Three Gorges Area, South China, and Their Ecological Implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 505: 150-166. https://doi.org/10.1016/j.palaeo.2018.05.039 |
| Chen, D. Z., Wang, J. G., Qing, H. R., et al., 2009. Hydrothermal Venting Activities in the Early Cambrian, South China: Petrological, Geochronological and Stable Isotopic Constraints. Chemical Geology, 258(3/4): 168-181. https://doi.org/10.1016/j.chemgeo.2008.10.016 |
| Chen, X., Ling, H. F., Vance, D., et al., 2015. Rise to Modern Levels of Ocean Oxygenation Coincided with the Cambrian Radiation of Animals. Nature Communications, 6(1): 7142. https://doi.org/10.1038/ncomms8142 |
| Ciglenečki, I., Ćosović, B., Vojvodić, V., et al., 2000. The Role of Reduced Sulfur Species in the Coalescence of Polysaccharides in the Adriatic Sea. Marine Chemistry, 71(3/4): 233-249. https://doi.org/10.1016/s0304-4203(00)00051-7 |
| Cremonese, L., Shields-Zhou, G. A., Struck, U., et al., 2014. Nitrogen and Organic Carbon Isotope Stratigraphy of the Yangtze Platform during the Ediacaran-Cambrian Transition in South China. Palaeogeography, Palaeoclimatology, Palaeoecology, 398: 165-186. https://doi.org/10.1016/j.palaeo.2013.12.016 |
| Demaison, G. J., Moore, G. T., 1980. Anoxic Environments and Oil Source Bed Genesis. Organic Geochemistry, 2(1): 9-31. https://doi.org/10.1016/0146-6380(80)90017-0 |
| Dennett, M. R., 2002. Video Plankton Recorder Reveals High Abundances of Colonial Radiolaria in Surface Waters of the Central North Pacific. Journal of Plankton Research, 24(8): 797-805. https://doi.org/10.1093/plankt/24.8.797 |
| Ding, Z. J., Bourven, I., Guibaud, G., et al., 2015. Role of Extracellular Polymeric Substances (EPS) Production in Bioaggregation: Application to Wastewater Treatment. Applied Microbiology and Biotechnology, 99(23): 9883-9905. https://doi.org/10.1007/s00253-015-6964-8 |
| Dong, L., Shen, B., Lee, C. T. A., et al., 2015. Germanium/Silicon of the Ediacaran-Cambrian Laobao Cherts: Implications for the Bedded Chert Formation and Paleoenvironment Interpretations. Geochemistry, Geophysics, Geosystems, 16(3): 751-763. https://doi.org/10.1002/2014gc005595 |
| Du, R. L., Tian, L. F., 1986. Macroalgae of Qingbaikouan Period in Yanshan Area. Hebei Science and Technology Press, Shijiazhuang. 114 (in Chinese) |
| Flemming, H. C., Neu, T. R., Wozniak, D. J., 2007. The EPS Matrix: The "House of Biofilm Cells". Journal of Bacteriology, 189(22): 7945-7947. https://doi.org/10.1128/jb.00858-07 |
| Gelin, F., Volkman, J. K., Largeau, C., et al., 1999. Distribution of Aliphatic, Nonhydrolyzable Biopolymers in Marine Microalgae. Organic Geochemistry, 30(2/3): 147-159. https://doi.org/10.1016/s0146-6380(98)00206-x |
| Gendron-Badou, A., Coradin, T., Maquet, J., et al., 2003. Spectroscopic Characterization of Biogenic Silica. Journal of Non-Crystalline Solids, 316(2/3): 331-337. https://doi.org/10.1016/s0022-3093(02)01634-4 |
| Graz, Y., Di-Giovanni, C., Copard, Y., et al., 2010. Quantitative Palynofacies Analysis as a New Tool to Study Transfers of Fossil Organic Matter in Recent Terrestrial Environments. International Journal of Coal Geology, 84(1): 49-62. https://doi.org/10.1016/j.coal.2010.08.006 |
| Guilbaud, R., Slater, B. J., Poulton, S. W., et al., 2018. Oxygen Minimum Zones in the Early Cambrian Ocean. Geochemical Perspectives Letters, 6: 33-38. https://doi.org/10.17863/cam.22469 |
| Guo, J. F., 2009. Yanjiahe Biota from the Early Cambrian of Yichang, Hubei, China: [Dissertation]. Northwest University, Chongqing. 166 (in Chinese with English Abstract) |
| Guo, J. F., Li, Y., Shu, D., 2010. Fossil Macroscopic Algae from the Yanjiahe Formation, Terreneuvian of the Three Gorges Area, South China. Acta Palaeonotologica Sinica, 49(3): 144-149 (in Chinese with English Abstract) http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=gswxb201003005 |
| Guo, J. F., Li, Y., Li, G. X., 2014. Small Shelly Fossils from the Early Cambrian Yanjiahe Formation, Yichang, Hubei, China. Gondwana Research, 25(3): 999-1007. https://doi.org/10.1016/j.gr.2013.03.007 |
| Guo, Q. J., Strauss, H., Liu, C. Q., et al., 2007a. Carbon Isotopic Evolution of the Terminal Neoproterozoic and Early Cambrian: Evidence from the Yangtze Platform, South China. Palaeogeography, Palaeoclimatology, Palaeoecology, 254(1/2): 140-157. https://doi.org/10.1016/j.palaeo.2007.03.014 |
| Guo, Q. J., Shields, G. A., Liu, C. Q., et al., 2007b. Trace Element Chemostratigraphy of Two Ediacaran-Cambrian Successions in South China: Implications for Organosedimentary Metal Enrichment and Silicification in the Early Cambrian. Palaeogeography, Palaeoclimatology, Palaeoecology, 254(1/2): 194-216. https://doi.org/10.1016/j.palaeo.2007.03.016 |
| Guo, Q. J., Strauss, H., Zhu, M. Y., et al., 2013. High Resolution Organic Carbon Isotope Stratigraphy from a Slope to Basinal Setting on the Yangtze Platform, South China: Implications for the Ediacaran-Cambrian Transition. Precambrian Research, 225: 209-217. https://doi.org/10.1016/j.precamres.2011.10.003 |
| Haq, B. U., Schutter, S. R., 2008. A Chronology of Paleozoic Sea-Level Changes. Science, 322(5898): 64-68. https://doi.org/10.1126/science.1161648 |
| Hartnett, H. E., Keil, R. G., Hedges, J. I., et al., 1998. Influence of Oxygen Exposure Time on Organic Carbon Preservation in Continental Margin Sediments. Nature, 391(6667): 572-575. https://doi.org/10.1038/35351 |
| Hatch, J. R., Leventhal, J. S., 1992. Relationship between Inferred Redox Potential of the Depositional Environment and Geochemistry of the Upper Pennsylvanian (Missourian) Stark Shale Member of the Dennis Limestone, Wabaunsee County, Kansas, U.S.A.. Chemical Geology, 99(1/2/3): 65-82. https://doi.org/10.1016/0009-2541(92)90031-y |
| Hu, J., Zhang, S. T., Zhang, G. Z., et al., 2018. Geochemistry and Tectonic Setting of the Eshan Granites in the Southwestern Margin of the Yangtze Plate, Yunnan. Journal of Earth Science, 29(1): 130-143. https://doi.org/10.1007/s12583-017-0747-3 |
| Hu, J., 2008. The Cherty Microbolite in the Deeper Water Facies during the Precambrian-Cambrian Transitional Period in Northeast Guangxi Province, China. Acta Micropalaeontologica Sinia, 25(3): 291-305 (in Chinese with English Abstract) http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=wtgswxb200803008 |
| Hu, L., Zhu, Y. M., Chen, S. B., et al., 2012. Resource Potential Analysis of Shale Gas in Lower Cambrian Qiongzhusi Formation in Middle & Upper Yangtze Region. Journal of China Coal Society, 37(11): 1871-1877 http://d.old.wanfangdata.com.cn/Periodical/mtxb201211018 |
| Huc, A. Y., Bertrand, P., Stow, D. A. V., 2000. Depositional Processes of Source Rocks in Deep Offshore Settings; Quaternary Analogs. Annual Meeting Expanded Abstracts-American Association of Petroleum Geologists, 70 http://cn.bing.com/academic/profile?id=f700bfae28d166f8d02b750afc7df6bb&encoded=0&v=paper_preview&mkt=zh-cn |
| Jiang, G. Q., Wang, X. Q., Shi, X. Y., et al., 2012. The Origin of Decoupled Carbonate and Organic Carbon Isotope Signatures in the Early Cambrian (ca. 542-520 Ma) Yangtze Platform. Earth and Planetary Science Letters, 317-318: 96-110. https://doi.org/10.1016/j.epsl.2011.11.018 |
| Jiang, X. F., Peng, S. B., Kusky, T. M., et al., 2018. Petrogenesis and Geotectonic Significance of Early-Neoproterozoic Olivine-Gabbro within the Yangtze Craton: Constrains from the Mineral Composition, U-Pb Age and Hf Isotopes of Zircons. Journal of Earth Science, 29(1): 93-102. https://doi.org/10.1007/s12583-018-0821-5 |
| Jin, C. S., Li, C., Algeo, T. J., et al., 2016. A Highly Redox-Heterogeneous Ocean in South China during the Early Cambrian (~529-514 Ma): Implications for Biota-Environment Co-Evolution. Earth and Planetary Science Letters, 441: 38-51. https://doi.org/10.1016/j.epsl.2016.02.019 |
| Jones, B., Manning, D. A. C., 1994. Comparison of Geochemical Indices Used for the Interpretation of Palaeoredox Conditions in Ancient Mudstones. Chemical Geology, 111(1/2/3/4): 111-129. https://doi.org/10.1016/0009-2541(94)90085-x |
| Jack, C. R. Jr, Knopman, D. S., Jagust, W. J., et al., 2010. Hypothetical Model of Dynamic Biomarkers of the Alzheimer's Pathological Cascade. The Lancet Neurology, 9(1): 119-128. https://doi.org/10.1016/s1474-4422(09)70299-6 |
| Kennedy, M. J., 2002. Mineral Surface Control of Organic Carbon in Black Shale. Science, 295(5555): 657-660. https://doi.org/10.1126/science.1066611 |
| Lee, C., Wakeham, S. G., 1992. Organic Matter in the Water Column: Future Research Challenges. Marine Chemistry, 39(1/2/3): 95-118. https://doi.org/10.1016/0304-4203(92)90097-t |
| Lebrato, M., Pahlow, M., Oschlies, A., et al., 2011. Depth Attenuation of Organic Matter Export Associated with Jelly Falls. Limnology and Oceanography, 56(5): 1917-1928. https://doi.org/10.4319/lo.2011.56.5.1917 |
| Lebrato, M., Mendes, P. D. J., Steinberg, D. K., et al., 2013. Jelly Biomass Sinking Speed Reveals a Fast Carbon Export Mechanism. Limnology and Oceanography, 58(3): 1113-1122. https://doi.org/10.4319/lo.2013.58.3.1113 |
| Lei, Y., Servais, T., Feng, Q. L., et al., 2012. The Spatial (Nearshore-Offshore) Distribution of Latest Permian Phytoplankton from the Yangtze Block, South China. Palaeogeography, Palaeoclimatology, Palaeoecology, 363-364: 151-162. https://doi.org/10.1016/j.palaeo.2012.09.010 |
| Li, J., Xie, X., Lin, Z., 2009. Organic Matter Enrichment of the Dalong Formation in Guangyuan Area of the Sichuan Basin. Geological Science and Technology Information, 28(2): 98-103 (in Chinese with English Abstract) http://cn.bing.com/academic/profile?id=1cc5a5332b6d82120086df84dcb11a86&encoded=0&v=paper_preview&mkt=zh-cn |
| Li, J., He, D., 2014. Palaeogeography and Tectonic-Depositional Environment Evolution of the Cambrian in Sichuan Basin and Adjacent Areas. Journal of Palaeogeography, 16(4): 441-460 (in Chinese with English Abstract) http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=gdlxb201404002 |
| Li, J. G., Batten, D. J., 2005. Palynofacies: Principles and Methods. Acta Palaeontologica Sinica, 44(1): 138-156 (in Chinese with English Abstract) http://d.old.wanfangdata.com.cn/NSTLQK/NSTL_QKJJ0231733855/ |
| Li, Z. Q., 2015. The Distribution and Influencing Factors of Terrestrial Organic Matter in the Typical Systems: [Dissertation]. East China Normal University, Shanghai. 186 (in Chinese with English Abstract) |
| Little, S. H., Vance, D., Lyons, T. W., et al., 2015. Controls on Trace Metal Authigenic Enrichment in Reducing Sediments: Insights from Modern Oxygen-Deficient Settings. American Journal of Science, 315(2): 77-119. https://doi.org/10.2475/02.2015.01 |
| Lu, L., Qin, J. Z., Shen, B. J., et al., 2018. Biogenetic Evidence of Siliceous Shale in the Wufeng Formation-Longmaxi Formation in the Upper Yangtze Area and Its Relationship with Shale Gas Enrichment. Journal of Geoscience, 4: 226-236 (in Chinese with English Abstract) |
| Luo, H. L., Jiang, Z. W., Tang, L. D., 1994. Stratotype Section for Lower Cambrian Stages in China. Yunnan Science and Technology Press, Kunming. 183 (in Chinese with English Abstract) |
| Lyle, M., Murray, D. W., Finney, B. P., et al., 1988. The Record of Late Pleistocene Biogenic Sedimentation in the Eastern Tropical Pacific Ocean. Paleoceanography, 3(1): 39-59. https://doi.org/10.1029/pa003i001p00039 |
| Ma, Q. F., Feng, Q. L., Cao, W. C., et al., 2019. Radiolarian Fauna from the Chiungchussuan Shuijingtuo Formation (Cambrian Series 2) in Western Hubei Province, South China. Science China Earth Sciences, 62: 1-14. https://doi.org/10.1007/s11430-018-9335-0 |
| Maliva, R. G., Knoll, A. H., Siever, R., 1989. Secular Change in Chert Distribution: A Reflection of Evolving Biological Participation in the Silica Cycle. Palaios, 4(6): 519-532. https://doi.org/10.2307/3514743 |
| Mecozzi, M., Acquistucci, R., Di Noto, V., et al., 2001. Characterization of Mucilage Aggregates in Adriatic and Tyrrhenian Sea: Structure Similarities between Mucilage Samples and the Insoluble Fractions of Marine Humic Substance. Chemosphere, 44(4): 709-720. https://doi.org/10.1016/s0045-6535(00)00375-1 |
| Meyers, S. R., Sageman, B. B., Lyons, T. W., 2005. Organic Carbon Burial Rate and the Molybdenum Proxy: Theoretical Framework and Application to Cenomanian-Turonian Oceanic Anoxic Event 2. Paleoceanography, 20(2): 169-189. https://doi.org/10.1029/2004pa001068 |
| Miller, K. G., 2005. The Phanerozoic Record of Global Sea-Level Change. Science, 310(5752): 1293-1298. https://doi.org/10.1126/science.1116412 |
| Mo, X., 2012. Study of Stratigraphic Classification and the Variance of Sedimentary System of Cambrian Stratum in Guangyuan Area: [Dissertation]. Chengdu University of Technology, Chengdu. 69 (in Chinese with English Abstract) |
| Moczydłowska, M., Zang, W. L., 2006. The Early Cambrian Acritarch Skiagia and Its Significance for Global Correlation. Palaeoworld, 15(3/4): 328-347. https://doi.org/10.1016/j.palwor.2006.10.003 |
| Moczydłowska, M., Willman, S., 2009. Ultrastructure of Cell Walls in Ancient Microfossils as a Proxy to Their Biological Affinities. Precambrian Research, 173(1/2/3/4): 27-38. https://doi.org/10.1016/j.precamres.2009.02.006 |
| Moczydłowska, M., 2011. The Early Cambrian Phytoplankton Radiation: Acritarch Evidence from the Lükati Formation, Estonia. Palynology, 35(1): 103-145. https://doi.org/10.1080/01916122.2011.552563 |
| Obeid, W., Salmon, E., Lewan, M. D., et al., 2015. Hydrous Pyrolysis of Scenedesmus Algae and Algaenan-Like Residue. Organic Geochemistry, 85: 89-101. https://doi.org/10.1016/j.orggeochem.2015.04.001 |
| Parrish, J. T., 1982. Upwelling and Petroleum Source Beds, with Reference to Palaeozoic, Deep Sea Research Part B. AAPG Bulletin, 66: 750-774. https://doi.org/10.1306/03b5a30e-16d1-11d7-8645000102c1865d |
| Pedersen, T. F., Calvert, S. E., 1990. Anoxia versus Productivity: What Controls the Formation of Organic-Carbon-Rich Sediments and Sedimentary Rocks? (1). AAPG Bulletin, 74(4): 454-466. https://doi.org/10.1306/0c9b232b-1710-11d7-8645000102c1865d |
| Piper, D. Z., Perkins, R. B., 2004. A Modern vs. Permian Black Shale—The Hydrography, Primary Productivity, and Water-Column Chemistry of Deposition. Chemical Geology, 206(3/4): 177-197. https://doi.org/10.1016/j.chemgeo.2003.12.006 |
| Poulton, S. W., Fralick, P. W., Canfield, D. E., 2010. Spatial Variability in Oceanic Redox Structure 1.8 Billion Years Ago. Nature Geoscience, 3(7): 486-490. https://doi.org/10.1038/ngeo889 |
| Racki, G., Cordey, F., 2000. Radiolarian Palaeoecology and Radiolarites: Is the Present the Key to the Past?. Earth-Science Reviews, 52(1/2/3): 83-120. https://doi.org/10.1016/s0012-8252(00)00024-6 |
| Robinson, D. H., Sullivan, C. W., 1987. How do Diatoms Make Silicon Biominerals?. Trends in Biochemical Sciences, 12: 151-154. https://doi.org/10.1016/0968-0004(87)90072-7 |
| Ross, D. J. K., Marc Bustin, R., 2009. The Importance of Shale Composition and Pore Structure upon Gas Storage Potential of Shale Gas Reservoirs. Marine and Petroleum Geology, 26(6): 916-927. https://doi.org/10.1016/j.marpetgeo.2008.06.004 |
| Sharma, M., Mishra, S., Dutta, S., et al., 2009. On the Affinity of Chuaria-Tawuia Complex: A Multidisciplinary Study. Precambrian Research, 173(1/2/3/4): 123-136. https://doi.org/10.1016/j.precamres.2009.04.003 |
| Shen, J., Schoepfer, S. D., Feng, Q. L., et al., 2015. Marine Productivity Changes during the End-Permian Crisis and Early Triassic Recovery. Earth-Science Reviews, 149: 136-162. https://doi.org/10.1016/j.earscirev.2014.11.002 |
| Steiner, M., Zhu, M., Weber, B., et al., 2001. The Lower Cambrian of Eastern Yunnan: Trilobite-Based Biostratigraphy and Related Faunas. Acta Palaeont Sinica, 40: 63-79 (in Chinese with English Abstract) https://www.researchgate.net/publication/236610439_The_Lower_Cambrian_of_Eastern_Yunnan_Trilobite-based_biostratigraphy_and_related_faunas |
| Steiner, M., Li, G. X., Qian, Y., et al., 2004. Lower Cambrian Small Shelly Fossils of Northern Sichuan and Southern Shaanxi (China), and Their Biostratigraphic Importance. Geobios, 37(2): 259-275. https://doi.org/10.1016/j.geobios.2003.08.001 |
| Taylor, S. R., McLennan, S. M., 1985. The Continental Crust: Its Composition and Evolution, Blackwell Scientific Publications, Oxford |
| Tian, W., Pan, L., Jiang, L., 2001. A Discussion on "Paleo-Island of Central Hubei" in Early Stage of Lower Cambrian Epoch. Hubei Geology & Mineral Resources, 15(4): 7-11 (in Chinese with English Abstract) http://dict.cnki.net/h_242256000.html |
| Traverse, A., 2007. Differential Sorting of Palynomorphs into Sediments: Palynofacies, Palynodebris, Discordant Palynomorphs. Paleopalynology, 579: 275-287. https://doi.org/10.1007/978-1-4020-5610-9_18 |
| Tribovillard, N., Algeo, T. J., Lyons, T., et al., 2006. Trace Metals as Paleoredox and Paleoproductivity Proxies: An Update. Chemical Geology, 232(1/2): 12-32. https://doi.org/10.1016/j.chemgeo.2006.02.012 |
| Tucker, M. E., 1992. The Precambrian-Cambrian Boundary: Seawater Chemistry, Ocean Circulation and Nutrient Supply in Metazoan Evolution, Extinction and Biomineralization. Journal of the Geological Society, 149(4): 655-668. https://doi.org/10.1144/gsjgs.149.4.0655 |
| Turgeon, S., Brumsack, H. J., 2006. Anoxic vs Dysoxic Events Reflected in Sediment Geochemistry during the Cenomanian-Turonian Boundary Event (Cretaceous) in the Umbria-Marche Basin of Central Italy. Chemical Geology, 234(3/4): 321-339. https://doi.org/10.1016/j.chemgeo.2006.05.008 |
| Tyson, R. V., 1987. The Genesis and Palynofacies Characteristics of Marine Petroleum Source Rocks. Geological Society, London, Special Publications, 26(1): 47-67. https://doi.org/10.1144/gsl.sp.1987.026.01.03 |
| Tyson, R. V., Pearson, T. H., 1991. Modern and Ancient Continental Shelf Anoxia: An Overview. Geological Society, London, Special Publications, 58(1): 1-24. https://doi.org/10.1144/gsl.sp.1991.058.01.01 |
| Tyson, R. V., 2005. The "Productivity versus Preservation" Controversy; Cause, Flaws, Andresolution. In: Harris, N. B., ed., Deposition of |
| Organic-Carbon-Rich Sediments: Models, Mechanisms, and Consequences. Society for Sedimentary Geology Special Publication, 82: 17-33. https: //doi.org/10.2110/pec.05.82.0017 |
| van de Velde, S., 2018. Electron Shuttling and Elemental Cycling in the Seafloor: [Dissertation]. Vrije Universiteit Brussel/Universiteit Antwerpen, Brussel/Antwerpen. 352 |
| van de Velde, S., Mills, B. J. W., Meysman, F. J. R., et al., 2018. Early Palaeozoic Ocean Anoxia and Global Warming Driven by the Evolution of Shallow Burrowing. Nature Communications, 9(1): 2554-2564. https://doi.org/10.1038/s41467-018-04973-4 |
| Vandenbroucke, M., Largeau, C., 2007. Kerogen Origin, Evolution and Structure. Organic Geochemistry, 38(5): 719-833. https://doi.org/10.1016/j.orggeochem.2007.01.001 |
| Verdugo, P., Alldredge, A. L., Azam, F., et al., 2004. The Oceanic Gel Phase: A Bridge in the DOM-POM Continuum. Marine Chemistry, 92(1/2/3/4): 67-85. https://doi.org/10.1016/j.marchem.2004.06.017 |
| Verdugo, P., Santschi, P. H., 2010. Polymer Dynamics of DOC Networks and Gel Formation in Seawater. Deep Sea Research Part II: Topical Studies in Oceanography, 57(16): 1486-1493. https://doi.org/10.1016/j.dsr2.2010.03.002 |
| Verlaan, P. A., 2008. The Role of Primary-Producer-Mediated Organic Complexation in Regional Variation in the Supply of Mn, Fe, Co, Cu, Ni and Zn to Oceanic, Non-Hydrothermal Ferromanganese Crusts and Nodules. Marine Georesources & Geotechnology, 26(4): 214-230. https://doi.org/10.1080/10641190802459704 |
| Volcani, B. E., Simpson, T. L., Volcani, B. E., 1981. Silicon and Siliceous Structures in Biological Systems. 157-200. https://doi.org/10.1007/978-1-4612-5944-2_2 |
| Wang, J. G., Chen, D. Z., Yan, D. T., et al., 2012. Evolution from an Anoxic to Oxic Deep Ocean during the Ediacaran-Cambrian Transition and Implications for Bioradiation. Chemical Geology, 306-307: 129-138. https://doi.org/10.1016/j.chemgeo.2012.03.005 |
| Wang, J., Li, X., Huang, W., 2014. The Shale Gas Exploration Prospect Assesses of the Niutitang Formation in Western Hubei and Hunan-Eastern Chongqing. Geological Science and Technology Information, 33: 98-103 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-DZKQ201404016.htm |
| Wu, W., Zhu, M. Y., Steiner, M., 2014. Composition and Tiering of the Cambrian Sponge Communities. Palaeogeography, Palaeoclimatology, Palaeoecology, 398: 86-96. https://doi.org/10.1016/j.palaeo.2013.08.003 |
| Xia, M. L., Wen, L., Wang, Y. G., et al., 2010. High Quality Source Rocks in Trough Facies of Upper Permian Dalong Formation, Sichuan Basin. Petroleum Exploration and Development, 37(6): 654-662. https://doi.org/10.1016/s1876-3804(11)60002-5 |
| Xiang, Y., Feng, Q. L., Shen, J., et al., 2013. Changhsingian Radiolarian Fauna from Anshun of Guizhou, and Its Relationship to TOC and Paleo-Productivity. Science China Earth Sciences, 56(8): 1334-1342. https://doi.org/10.1007/s11430-013-4615-4 |
| Xiao, S. H., Hu, J., Yuan, X. L., et al., 2005. Articulated Sponges from the Lower Cambrian Hetang Formation in Southern Anhui, South China: Their Age and Implications for the Early Evolution of Sponges. Palaeogeography, Palaeoclimatology, Palaeoecology, 220(1/2): 89-117. https://doi.org/10.1016/j.palaeo.2002.02.001 |
| Xing, Y., 1982. Microflora of the Sinian System and Lower Cambrian near Kunming, Yunnan and Its Stratigraphical Significance. Acta Geologica Sinica, 56(1): 42-50 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-DZXE198201003.htm |
| Xu, L. G., Lehmann, B., Mao, J. W., et al., 2012. Mo Isotope and Trace Element Patterns of Lower Cambrian Black Shales in South China: Multi-Proxy Constraints on the Paleoenvironment. Chemical Geology, 318-319: 45-59. https://doi.org/10.1016/j.chemgeo.2012.05.016 |
| Yamamoto, K., 1987. Geochemical Characteristics and Depositional Environments of Cherts and Associated Rocks in the Franciscan and Shimanto Terranes. Sedimentary Geology, 52(1/2): 65-108. https://doi.org/10.1016/0037-0738(87)90017-0 |
| Yang, A. H., Zhu, M. Y., Zhang, J. M., et al., 2003. Early Cambrian Eodiscoid Trilobites of the Yangtze Platform and Their Stratigraphic Implications. Progress in Natural Science, 13(11): 861-866. https://doi.org/10.1080/10020070312331344560 |
| Yang, R. D., Zhao, Y. L., Guo, Q. J., 1999. Algae and Acritarchs and Their Palaeooceanographic Significance from the Early Cambrian Black Shale in Guizhou, China. Acta Palaeonto Logica Sinica, 38: 154-160 (in Chinese with English Abstract) http://cn.bing.com/academic/profile?id=14e5f5a5bec0e25a041f5cdac39a938a&encoded=0&v=paper_preview&mkt=zh-cn |
| Yin, F., Xue, X., 2001. Early Radiation of Acritarch and Its Significance. Journal of Northwest University (Natural Science Edition), 31(5): 409-411 (in Chinese with English Abstract) http://cn.bing.com/academic/profile?id=8beb93c975527cad79534b17dc670274&encoded=0&v=paper_preview&mkt=zh-cn |
| Yin, L., 1987. New Data of Microfossils from Precambrian-Cambrian Cherts in Ningqiang, South Shaanxi. Acta Palaeontologica Sinica, 26(2): 187-195 (in Chinese with English Abstract) |
| Yin, L., 2006. Acritarch Study in China. Science Press, Beijing. 222 (in Chinese) |
| Yuan, X. L., Xiao, S. H., Parsley, R. L., et al., 2002. Towering Sponges in an Early Cambrian Lagerstätte: Disparity between Nonbilaterian and Bilaterian Epifaunal Tierers at the Neoproterozoic-Cambrian Transition. Geology, 30(4): 363-366. https://doi.org/10.1130/0091-7613(2002)030 < 0363:tsiaec > 2.0.co; 2 doi: 10.1130/0091-7613(2002)030<0363:tsiaec>2.0.co;2 |
| Zhang, L., Danelian, T., Feng, Q. L., et al., 2013. On the Lower Cambrian Biotic and Geochemical Record of the Hetang Formation (Yangtze Platform, South China): Evidence for Biogenic Silica and Possible Presence of Radiolaria. Journal of Micropalaeontology, 32(2): 207-217. https://doi.org/10.1144/jmpaleo2013-003 |
| Zhang, L., 2014. Study on the Biota and Its Co-Evolution to the Paleoenvironment in the Early Cambrian of the Eastern Yangtze Gorges and Western Zhejiang, China: [Dissertation]. China University of Geosciences, Wuhan. 158 (in Chinese with English Abstract) |
| Zhang, S., Zhang, B., Bian, L., et al., 2007. The Accumulation of Red Algae from the Oil Shale of 800 Million Years Old Xiaoling Formation. Science in China Series D: Earth Science, 37(5): 636-643 (in Chinese with English Abstract) |
| Zhang, X. G., Pratt, B. R., 1994. New and Extraordinary Early Cambrian Sponge Spicule Assemblage from China. Geology, 22(1): 43-46. https://doi.org/10.1130/0091-7613(1994)022 < 0043:naeecs > 2.3.co; 2 doi: 10.1130/0091-7613(1994)022<0043:naeecs>2.3.co;2 |
| Zhang, L. W., Huang, J. H., Liang, Q., et al., 2007. Geological Characteristics and Ore Prospect of the Black Layers in the Doushantuo and Niutitang Formations in Guizhou Province. Acta Mineralogica Sinica, 27(1): 456-460 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTOTAL-KWXB2007Z1034.htm |
| Zhang, Y., Zheng, S., Gao, B., et al., 2017. Distribution Characteristics and Enrichment Factors of Organic Matter in Upper Permian Dalong Formation of Shangsi Section, Guangyuan, Sichuan Basin. Earth Science, 42(6): 1009-1025. https://doi.org/10.3799/dqkx.2017.534 (in Chinese with English Abstract) doi: 10.3799/dqkx.2017.534(inChinesewithEnglishAbstract) |
| Zhang, K., Feng, Q. L., 2019. Early Cambrian Radiolarians and Sponge Spicules from the Niujiaohe Formation in South China. Palaeoworld, 28(3): 234-242. https://doi.org/10.1016/j.palwor.2019.04.001. |
| Zhao, Y. L., Steiner, M., Yang, R. D., et al., 1999. Discovery and Significance of the Early Metazoan Biotas from the Lower Cambrian Niutitang Formation Zunyi, Guizhou, China. Acta Palaeontologica Sinica, 38(Suppl.): 139-153. https://doi.org/10.1108/jcom-04-2012-0030 |
| Zheng, H. L., Yang, X. L., Zhao, Y. L., et al., 2014. Stratigraphic Significance of Eodiscoids from the Niutitang Formation (Cambrian) in Jinsha County, Guizhou Province. Journal of Guizhou University (Natural Sciences), 31(1): 32-37 (in Chinese with English Abstract) http://en.cnki.com.cn/Article_en/CJFDTotal-GZDI201401008.htm |
| Zhou, C. M., Jiang, S. Y., 2009. Palaeoceanographic Redox Environments for the Lower Cambrian Hetang Formation in South China: Evidence from Pyrite Framboids, Redox Sensitive Trace Elements, and Sponge Biota Occurrence. Palaeogeography, Palaeoclimatology, Palaeoecology, 271(3/4): 279-286. https://doi.org/10.1016/j.palaeo.2008.10.024 |
| Zhu, X. J., Cai, J. G., Wang, X. J., et al., 2014. Effects of Organic Components on the Relationships between Specific Surface Areas and Organic Matter in Mudrocks. International Journal of Coal Geology, 133: 24-34. https://doi.org/10.1016/j.coal.2014.08.009 |
Figures(10) / Tables(1)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: