| Citation: | Jiyu Chen, Qiang Li, Qiufang He, Heinz C. Schröder, Zujun Lu, Daoxian Yuan. Influence of CO2/HCO3- on Microbial Communities in Two Karst Caves with High CO2. Journal of Earth Science, 2023, 34(1): 145-155. doi: 10.1007/s12583-020-1368-9

|
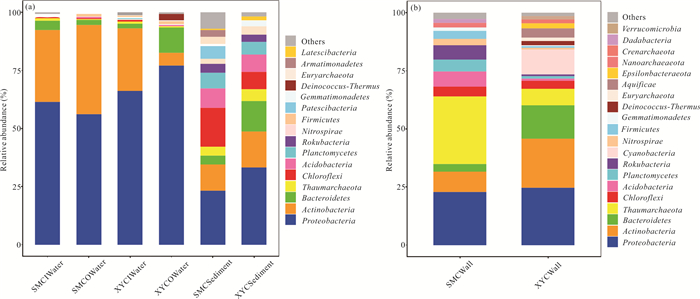
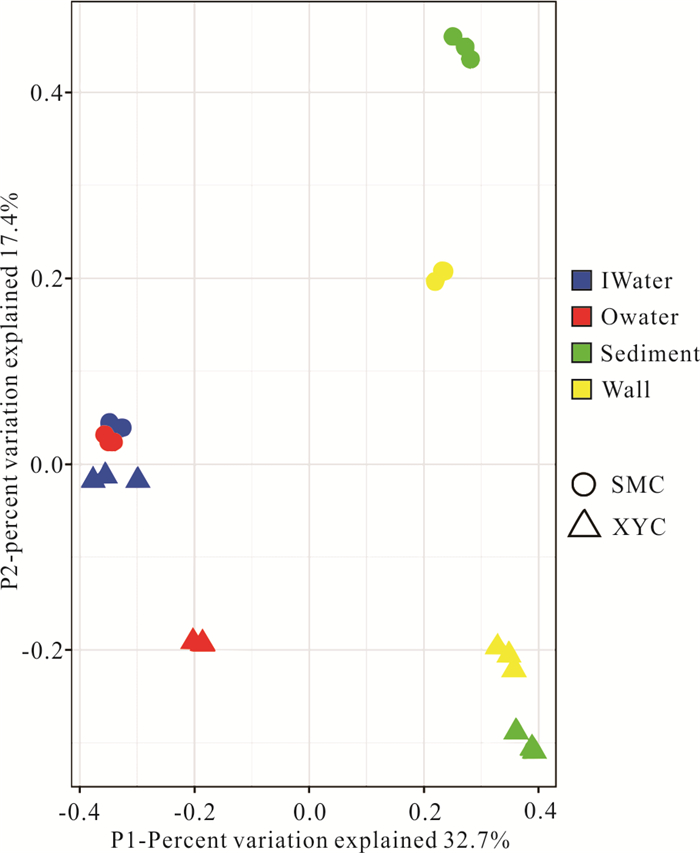
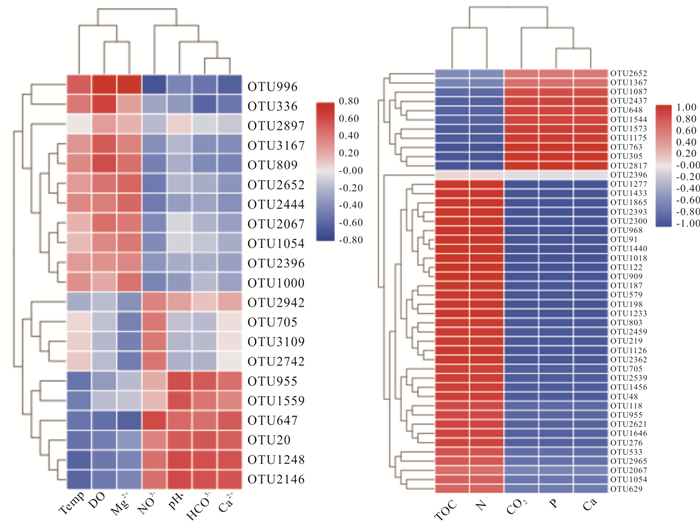
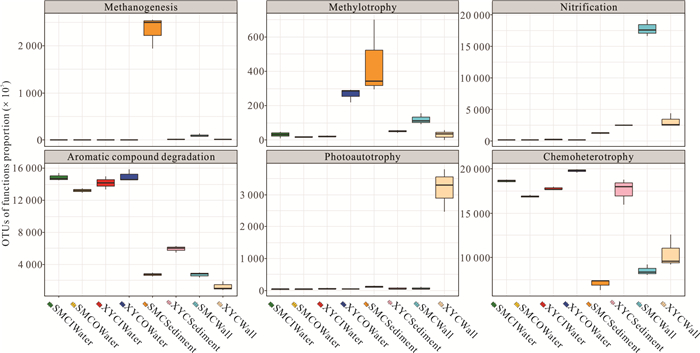
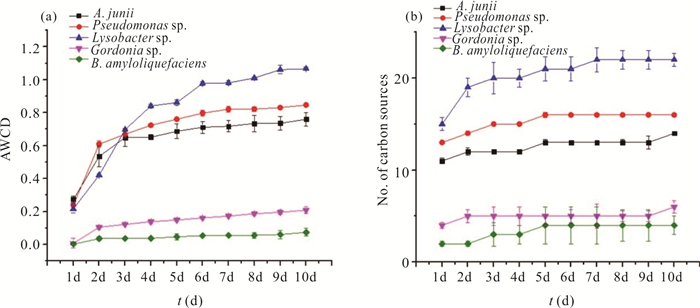
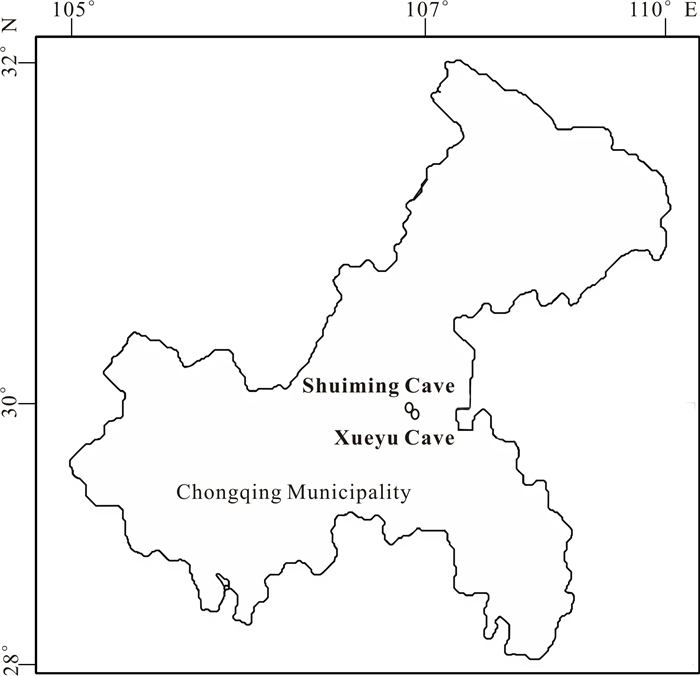
There is limited knowledge about microbial communities and their ecological functions in karst caves with high CO2 concentrations. Here, we studied the microbial community compositions and functions in Shuiming Cave ("SMC", CO2 concentration 3 303 ppm) and Xueyu Cave ("XYC", CO2 concentration 8 753 ppm) using Illumina MiSeq high-throughput sequencing in combination with BIOLOG test. The results showed that

| Anderson, M. J., Willis, T. J., 2003. Canonical Analysis of Principal Coordinates: A Useful Method of Constrained Ordination for Ecology. Ecology, 84(2): 511–525. https://doi.org/10.1890/0012-9658(2003)084[0511:caopca]2.0.co;2 |
| Baldini, J. U. L., Mcdermott, F., Hoffmann, D. L., et al., 2008. Very High-Frequency and Seasonal Cave Atmosphere PCO2 Variability: Implications for Stalagmite Growth and Oxygen Isotope-Based Paleoclimate Records. Earth and Planetary Science Letters, 272(1/2): 118–129. https://doi.org/10.1016/j.epsl.2008.04.031 |
| Barton, H. A., Taylor, M. R., Pace, N. R., 2004. Molecular Phylogenetic Analysis of a Bacterial Community in an Oligotrophic Cave Environment. Geomicrobiology Journal, 21(1): 11–20. https://doi.org/10.1080/01490450490253428 |
| Barton, H. A., Taylor, N. M., Kreate, M. P., et al., 2007. The Impact of Host Rock Geochemistry on Bacterial Community Structure in Oligotrophic Cave Environments. International Journal of Speleology, 36(2): 93–104. https://doi.org/10.5038/1827-806X.36.2.5 |
| Bauermeister, J., Ramette, A., Dattagupta, S., 2012. Repeatedly Evolved Host-Specific Ectosymbioses between Sulfur-Oxidizing Bacteria and Amphipods Living in a Cave Ecosystem. PLoS One, 7(11): e50254. https://doi.org/10.1371/journal.pone.0050254 |
| Bremner, J. M., 1960. Determination of Nitrogen in Soil by the Kjeldahl Method. The Journal of Agricultural Science, 55(1): 11–33. https://doi.org/10.1017/s0021859600021572 |
| Caporaso, J. G., Lauber, C. L., Walters, W. A., et al., 2012. Ultra-High-Throughput Microbial Community Analysis on the Illumina HiSeq and MiSeq Platforms. The ISME Journal, 6(8): 1621–1624. https://doi.org/10.1038/ismej.2012.8 |
| Chelius, M. K., Beresford, G., Horton, H., et al., 2009. Impacts of Alterations of Organic Inputs on the Bacterial Community within the Sediments of Wind Cave, South Dakota, USA. International Journal of Speleology, 38(1): 1–10. https://doi.org/10.5038/1827-806x.38.1.1 |
| Chen, F. H., Jia, J., Chen, J. H., et al., 2016. A Persistent Holocene Wetting Trend in Arid Central Asia, with Wettest Conditions in the Late Holocene, Revealed by Multi-Proxy Analyses of Loess-Paleosol Sequences in Xinjiang, China. Quaternary Science Reviews, 146: 134–146. https://doi.org/10.1016/j.quascirev.2016.06.002 |
| Chen, J. Y., Lu, Z. J., He, Q. F., et al., 2020. Characteristics of Culturable Bacterial Communities in Karst Caves with Different CO2 Concentrations―An Example from Xueyu Cave and Shuiming Cave in Chongqing. Carsologica Sinica, 39(2): 264–274. https://doi.org/10.11932/karst2020y20 (in Chinese with English Abstract) |
| de Mandal, S., Chatterjee, R., Kumar, N. S., 2017. Dominant Bacterial Phyla in Caves and Their Predicted Functional Roles in C and N Cycle. BMC Microbiology, 17(1): 90. https://doi.org/10.1186/s12866-017-1002-x |
| Deininger, M., Lippold, J., Abele, F., et al., 2016. Comparison of a Spatio-Temporal Speleothem-Based Reconstruction of Late Holocene Climate Variability to the Timing of Cultural Developments. EGU General Assembly Conference Abstracts |
| Deng, S. L., Liu, F. R., Zhang, Y. M., 2016. Effects of Elevated Temperature and Doubling of CO2 Concentration on the Soil Microbial Community Structure in the Subalpine Coniferous Forest of Western Sichuan, China. Chinese Journal of Applied and Environmental Biology, 22(1): 20–26 https://doi.org/10.3724/SP.J.1145.2015.06021 (in Chinese with English Abstract) |
| Dubbs, L. L., Whalen, S. C., 2010. Reduced Net Atmospheric CH4 Consumption is a Sustained Response to Elevated CO2 in a Temperate Forest. Biology and Fertility of Soils, 46(6): 597–606. https://doi.org/10.1007/s00374-010-0467-7 |
| Ebersberger, D., Niklaus, P. A., Kandeler, E., 2003. Long Term CO2 Enrichment Stimulates N-Mineralisation and Enzyme Activities in Calcareous Grassland. Soil Biology and Biochemistry, 35(7): 965–972. https://doi.org/10.1016/S0038-0717(03)00156-1 |
| Edgar, R. C., 2013. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nature Methods, 10(10): 996–998. https://doi.org/10.1038/nmeth.2604 |
| Edgar, R. C., Haas, B. J., Clemente, J. C., et al., 2011. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics, 27(16): 2194–2200. https://doi.org/10.1093/bioinformatics/btr381 |
| Fu, G., Zhang, H. R., Li, S. W., et al., 2019. A Meta-Analysis of the Effects of Warming and Elevated CO2 on Soil Microbes. Journal of Resources and Ecology, 10(1): 69–76. https://doi.org/10.5814/j.issn.1674-764x.2019.01.009 |
| Fuhrman, J. A., 2009. Microbial Community Structure and Its Functional Implications. Nature, 459(7244): 193–199. https://doi.org/10.1038/nature08058 |
| Gao, K. S., Yu, A. J., 2000. Influence of CO2, Light and Watering on Growth of Nostoc Flagelliforme Mats. Journal of Applied Phycology, 12(2): 185–189. https://doi.org/10.1023/A:1008123203409 |
| Garrity, G. M., Holt, J. G., Castenholz, R. W., et al., 2001. Phylum BVI. Chloroflexi Phy. Nov. . In: Boone, D. R., Castenholz, R. W., Garrity, G. M., eds., Bergey's Manual® of Systematic Bacteriology. Springer, New York. 427–446. https://doi.org/10.1007/978-0-387-21609-6_23 |
| Goldfarb, K. C., Karaoz, U., Hanson, C. A., et al., 2011. Differential Growth Responses of Soil Bacterial Taxa to Carbon Substrates of Varying Chemical Recalcitrance. Frontiers in Microbiology, 2: 94. https://doi.org/10.3389/fmicb.2011.00094 |
| Groth, I., Vettermann, R., Schuetze, B., et al., 1999. Actinomycetes in Karstic Caves of Northern Spain (Altamira and Tito Bustillo). Journal of Microbiological Methods, 36(1/2): 115–122. https://doi.org/10.1016/S0167-7012(99)00016-0 |
| He, M., Mei, C. F., Sun, G. P., et al., 2016. The Effects of Molecular Properties on Ready Biodegradation of Aromatic Compounds in the OECD 301B CO2 Evolution Test. Archives of Environmental Contamination and Toxicology, 71(1): 133–145. https://doi.org/10.1007/s00244-015-0236-6 |
| Hiraishi, A., Hoshino, Y., Satoh, T., 1991. Rhodoferax Fermentans Gen. Nov., Sp. Nov., a Phototrophic Purple Nonsulfur Bacterium Previously Referred to as the "Rhodocyclus Gelatinosus-Like" Group. Archives of Microbiology, 155(4): 330–336. https://doi.org/10.1007/BF00243451 |
| Hutchens, E., Radajewski, S., Dumont, M. G., et al., 2004. Analysis of Methanotrophic Bacteria in Movile Cave by Stable Isotope Probing. Environmental Microbiology, 6(2): 111–120. https://doi.org/10.1046/j.1462-2920.2003.00543.x |
| Jeong, S. Y., Kim, T. G., 2019. Development of a Novel Methanotrophic Process with the Helper Micro-Organism Hyphomicrobium Sp. NM3. Journal of Applied Microbiology, 126(2): 534–544. https://doi.org/10.1111/jam.14140 |
| Jiang, H. C., Dong, H. L., Zhang, G. X., et al., 2006. Microbial Diversity in Water and Sediment of Lake Chaka, an Athalassohaline Lake in Northwestern China. Applied and Environmental Microbiology, 72(6): 3832–3845. https://doi.org/10.1128/AEM.02869-05 |
| Könneke, M., Bernhard, A. E., de la Torre, J. R., et al., 2005. Isolation of an Autotrophic Ammonia-Oxidizing Marine Archaeon. Nature, 437(7058): 543–546. https://doi.org/10.1038/nature03911 |
| Kramer, C., Gleixner, G., 2008. Soil Organic Matter in Soil Depth Profiles: Distinct Carbon Preferences of Microbial Groups during Carbon Transformation. Soil Biology and Biochemistry, 40(2): 425–433. https://doi.org/10.1016/j.soilbio.2007.09.016 |
| Li, Q., Song, A., Peng, W. J., et al., 2017. Contribution of Aerobic Anoxygenic Phototrophic Bacteria to Total Organic Carbon Pool in Aquatic System of Subtropical Karst Catchments, Southwest China: Evidence from Hydrochemical and Microbiological Study. FEMS Microbiology Ecology, 93(6): fix065. https://doi.org/10.1093/femsec/fix065 |
| Liu, X. H., Song, X. S., Wang, Y., et al., 2015. Effects of drought and double CO2 in atmosphere on soil microbial biomass and activities. Jiangsu Agricultural Sciences, 43(12): 336–338. https://doi.org/10.15889/j.issn.1002-1302.2015.12.106 (in Chinese with English Abstract) |
| Liu, Y. C., Whitman, W. B., 2008. Metabolic, Phylogenetic, and Ecological Diversity of the Methanogenic Archaea. Annals of the New York Academy of Sciences, 1125(1): 171–189. https://doi.org/10.1196/annals.1419.019 |
| Louca, S., Parfrey, L. W., Doebeli, M., 2016. Decoupling Function and Taxonomy in the Global Ocean Microbiome. Science, 353(6305): 1272–1277. https://doi.org/10.1126/science.aaf4507 |
| Lü, X. F., He, Q. F., Wang, Z. J., et al., 2019. Calcium Carbonate Precipitation Mediated by Bacterial Carbonic Anhydrase in a Karst Cave: Crystal Morphology and Stable Isotopic Fractionation. Chemical Geology, 530: 119331. https://doi.org/10.1016/j.chemgeo.2019.119331 |
| Luo, W. Q., 2009. Analysing Ecological Data. Journal of Applied Statistics, 36(2): 233–234. https://doi.org/10.1080/02664760802340267 |
| Magoč, T., Salzberg, S. L., 2011. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics, 27(21): 2957–2963. https://doi.org/10.1093/bioinformatics/btr507 |
| McCann, K. S., 2000. The Diversity–Stability Debate. Nature, 405(6783): 228–233. https://doi.org/10.1038/35012234 |
| McDonough, L. K., Iverach, C. P., Beckmann, S., et al., 2016. Spatial Variability of Cave-Air Carbon Dioxide and Methane Concentrations and Isotopic Compositions in a Semi-Arid Karst Environment. Environmental Earth Sciences, 75(8): 700. https://doi.org/10.1007/s12665-016-5497-5 |
| Paerl, H. W., Gardner, W. S., Havens, K. E., et al., 2016. Mitigating Cyanobacterial Harmful Algal Blooms in Aquatic Ecosystems Impacted by Climate Change and Anthropogenic Nutrients. Harmful Algae, 54: 213–222. https://doi.org/10.1016/j.hal.2015.09.009 |
| Palmer, A. N., 2017. Geology of Mammoth Cave. In: Hobbs, H. III, Olson, R., Winkler, E., et al., eds., Mammoth Cave: Cave and Karst Systems of the World. Springer. https://doi.org/10.1007/978-3-319-53718-4_6 |
| Pearson, A., McNichol, A. P., Benitez-Nelson, B. C., et al., 2001. Origins of Lipid Biomarkers in Santa Monica Basin Surface Sediment: A Case Study Using Compound-Specific δ14C Analysis. Geochimica et Cosmochimica Acta, 65(18): 3123–3137. https://doi.org/10.1016/S0016-7037(01)00657-3 |
| Pedersen, K., 2000. Exploration of Deep Intraterrestrial Microbial Life: Current Perspectives. FEMS Microbiology Letters, 185(1): 9–16. https://doi.org/10.1016/S0378-1097(00)00061-6 |
| Pierce, S., Sjögersten, S., 2009. Effects of below Ground CO2 Emissions on Plant and Microbial Communities. Plant and Soil, 325(1): 197. https://doi.org/10.1007/s11104-009-9969-1 |
| Rothschild, L. J., Mancinelli, R. L., 2001. Life in Extreme Environments. Nature, 409(6823): 1092–1101. https://doi.org/10.1038/35059215 |
| Team, R. D. C., 2009. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Computing, 14: 12–21. https://doi.org/10.1890/0012-9658(2002)083[3097:CFHIWS]2.0.CO;2. |
| Tian, C. Q., Shao, K., 2013. Determination of Total Phosphorus in Soils and Stream Sediments by Microwave Digestion-Phosphorus Molybdenum Blue Spectrophotometry. Metallurgical Analysis, 33(12): 52–56. https://doi.org/10.13228/j.issn.1000-7571.2013.12.010 (in Chinese with English Abstract) |
| Vanfossen, A. L., Verhaart, M. R. A., Kengen, S. M. W., et al., 2009. Carbohydrate Utilization Patterns for the Extremely Thermophilic Bacterium Caldicellulosiruptor Saccharolyticus Reveal Broad Growth Substrate Preferences. Applied and Environmental Microbiology, 75(24): 7718–7724. https://doi.org/10.1128/AEM.01959-09 |
| Wang, F. X., Cao, J. H., Huang, J. F., 1998. Biokarst in Cave Twilight Zones. Carsologica Sinica, 17(1): 41–48.https://doi.org/CNKI:SUN:ZGYR.0.1998-01-005 (in Chinese with English Abstract) |
| Wang, J. F., Wang, Y. H., Song, X. S., et al., 2017. Elevated Atmospheric CO2 and Drought Affect Soil Microbial Community and Functional Diversity Associated with Glycine Max. Revista Brasileira De Ciência Do Solo, 41: e0160460. https://doi.org/10.1590/18069657rbcs20160460 |
| Willis, K. J., MacDonald, G. M., 2011. Long-Term Ecological Records and Their Relevance to Climate Change Predictions for a Warmer World. Annual Review of Ecology, Evolution, and Systematics, 42(1): 267–287. https://doi.org/10.1146/annurev-ecolsys-102209-144704 |
| Wong, C. I., Breecker, D. O., 2015. Advancements in the Use of Speleothems as Climate Archives. Quaternary Science Reviews, 127: 1–18. https://doi.org/10.1016/j.quascirev.2015.07.019 |
| Wuchter, C., Schouten, S., Boschker, H. T. S., et al., 2003. Bicarbonate Uptake by Marine Crenarchaeota. FEMS Microbiology Letters, 219(2): 203–207. https://doi.org/10.1016/S0378-1097(03)00060-0 |
| Xie, S. C., Wang, F. P., Yan, J. X., et al., 2016. Geobiological Processes during Critical Environmental Transitions in Earth History. Science & Technology Information, 14(21): 176–177. https://doi.org/10.3969/j.issn.1672-3791.2016.21.107 (in Chinese with English Abstract) |
| Yang, S. H., Zheng, Q. S., Yuan, M. T., et al., 2019. Long-Term Elevated CO2 Shifts Composition of Soil Microbial Communities in a Californian Annual Grassland, Reducing Growth and N Utilization Potentials. Science of the Total Environment, 652: 1474–1481. https://doi.org/10.1016/j.scitotenv.2018.10.353 |
| Yun, Y., Cheng, X. Y., Wang, W. Q., et al., 2018. Seasonal Variation of Bacterial Community and Their Functional Diversity in Drip Water from a Karst Cave. Chinese Science Bulletin, 63(36): 3932–3944, 3932. https://doi.org/10.1360/N972018-00627 (in Chinese with English Abstract) |
| Zhang, J., Gu, T., Zhou, Y., et al., 2012. Terrimonas Rubra Sp. Nov., Isolated from a Polluted Farmland Soil and Emended Description of the Genus Terrimonas. International Journal of Systematic and Evolutionary Microbiology, 62(Pt_11): 2593–2597. https://doi.org/10.1099/ijs.0.036079-0 |
| Zhang, L. M., He, J. Z., 2012. A Novel Archaeal Phylum: Thaumarchaeota—A Review. Acta Microbiologica Sinica, 52(4): 411–421. https://doi. org/http://ir.rcees.ac.cn/handle/311016/8332. |
| Zhang, Z. H., Li, X. N., Peng T., et al., 2004. New Records of Luminous Liverworts from the Karst Caves of Guangxi Province, P. R. China: Cyathodium Cavernarum Kunze and C. Smaragdium Schiffin ex Keissler (Cyathodiaceae, Hepaticae). Carsologica Sinica, 23(2): 154–157. https://doi.org/10.3969/j.issn.1001-4810.2004.02.011 (in Chinese with English Abstract) |
| Zhao, H. B., Xu, B. Q., Yao, T. D., et al., 2012. Deuterium Excess Record in a Southern Tibetan Ice Core and Its Potential Climatic Implications. Climate Dynamics, 38(9): 1791–1803. https://doi.org/10.1007/s00382-011-1161-7 |
| Zhao, J., Lu, W., Zhang, F. J., et al., 2014. Evaluation of CO2 Solubility-Trapping and Mineral-Trapping in Microbial-Mediated CO2-Brine-Sandstone Interaction. Marine Pollution Bulletin, 85(1): 78–85. https://doi.org/10.1016/j.marpolbul.2014.06.019 |
Figures(6) / Tables(2)
Copyright © 2013-2020 Journal of Earth Science 鄂ICP备15021562号-2
Tel: +86-27-67885075 Fax: +86-27-67885075 E-mail: xbb@cug.edu.cn
Address: Editorial Office of Journal, China University of Geosciences, Yujiashan, Wuhan, Hubei 430074, P. R. China
Supported by:
Beijing Renhe Information Technology Co. Ltd
E-mail:
info@rhhz.net



 DownLoad:
DownLoad: